- Гідрологія і Гідрометрія
- Господарське право
- Економіка будівництва
- Економіка природокористування
- Економічна теорія
- Земельне право
- Історія України
- Кримінально виконавче право
- Медична радіологія
- Методи аналізу
- Міжнародне приватне право
- Міжнародний маркетинг
- Основи екології
- Предмет Політологія
- Соціальне страхування
- Технічні засоби організації дорожнього руху
- Товарознавство продовольчих товарів
Тлумачний словник
Авто
Автоматизація
Архітектура
Астрономія
Аудит
Біологія
Будівництво
Бухгалтерія
Винахідництво
Виробництво
Військова справа
Генетика
Географія
Геологія
Господарство
Держава
Дім
Екологія
Економетрика
Економіка
Електроніка
Журналістика та ЗМІ
Зв'язок
Іноземні мови
Інформатика
Історія
Комп'ютери
Креслення
Кулінарія
Культура
Лексикологія
Література
Логіка
Маркетинг
Математика
Машинобудування
Медицина
Менеджмент
Метали і Зварювання
Механіка
Мистецтво
Музика
Населення
Освіта
Охорона безпеки життя
Охорона Праці
Педагогіка
Політика
Право
Програмування
Промисловість
Психологія
Радіо
Регилия
Соціологія
Спорт
Стандартизація
Технології
Торгівля
Туризм
Фізика
Фізіологія
Філософія
Фінанси
Хімія
Юриспунденкция
Властивості м’язового волокна. Механізми утворення та передачі збудження, скорочення скелетних м’язів.
У хребетних і людини три види м'язів: поперечнополосатиє м'язи скелета, поперечнополосата м'яз серця – міокард і гладкі м'язи, що утворюють стінки порожнистих внутрішніх органів і судин.
Анатомічної та функціональної одиницею скелетних м'язів є нейромоторная одиниця – руховий нейрон і иннервируемая їм група м'язових волокон. Імпульси, що посилаються мотонейроном, приводять в дію всі утворюючі її м'язові волокна.
Структурна організація м'язового волокна. М'язове волокно є багатоядерної структурою, оточеній мембраною і містить спеціалізований скорочувальний апарат – міофібрили (рис. 5. Крім цього, найважливішими компонентами м'язового волокна є мітохондрії, системи поздовжніх трубочок – саркоплазматична мережа (ретикулум), і система поперечних трубочок – Т-система. Функціональною одиницею скоротливого апарату м'язової клітини є саркомер; із саркомерів складається міофібрила. Саркомери відокремлюються один від одного Z-пластинками. Саркомери в міофібрилі розташовані послідовно, тому скорочення саркомерів викликає скорочення міофібрили і загальне вкорочення м'язового волокна.
В період відносного спокою скелетні м'язи повністю не розслабляються і зберігають помірний ступінь напруги, тобто м'язовий тонус.
Основні функції м'язової тканини:
- рухова – забезпечення руху;
- статична – забезпечення фіксації, в тому числі і в певній позі;
- рецепторна – у м'язах є рецептори, що дозволяють сприймати власні рухи;
- депонує – в м'язах запасаються вода і деякі поживні речовини.
Фізіологічні властивості скелетних м'язів:
1. Збудливість. Нижче, ніж збудливість нервової тканини. Збудження поширюється уздовж м'язового волокна.
2. Провідність. Менше провідності нервової тканини.
3. Рефрактерний період м'язової тканини більш тривалий, ніж нервової тканини.
4. Лабільність м'язової тканини значно нижче, ніж нервової.
5. Скорочення – здатність м'язового волокна змінювати свою довжину і ступінь напруження у відповідь на подразнення порогової сили.
При ізотонічному скороченні змінюється довжина м'язового волокна без зміни тонусу. При ізометричному скороченні зростає напруження м'язового волокна без зміни його довжини.
В залежності від умов стимуляції і функціонального стану м'яза може виникнути одиночне, злите (тетаніческе) скорочення або контрактура м'яза.

Рисунок 5– Структура м'язового волокна
Одиночне м'язове скорочення. При подразненні м'язи одиночним імпульсом струму виникає одиночне м'язове скорочення.
Амплітуда одиночного скорочення м'яза залежить від кількості міофібрил, що скоротились в цей момент. Збудливість окремих груп волокон різна, тому порогова сила струму викликає скорочення лише найбільш збудливих м'язових волокон. Амплітуда такого скорочення мінімальна. При збільшенні сили дратівної струму в процес збудження утягуються і менш збудливі групи м'язових волокон; амплітуда скорочень підсумовується і росте до тих пір, поки в м'язі не залишиться волокон, не охоплених процесом порушення. У цьому випадку реєструється максимальна амплітуда скорочення, яка не збільшується, незважаючи на подальше наростання сили дратівної струму.
Тетаніческе скорочення. У природних умовах до м'язових волокон надходять не поодинокі, а ряд нервових імпульсів, на які м'яз відповідає тривалим, тетаніческое скорочення, або тетанус. До тетаніческого скорочення здатні тільки скелетні м'язи. Гладкі м'язи і поперечнополосатий м'яз серця не здатні до тетаніческого скорочення через тривалу рефрактерного періоду.
Тетанус виникає внаслідок сумації одиночних м'язових скорочень. Щоб виник тетанус, необхідно дію повторних подразнень (або нервових імпульсів) на м'яз ще до того, як закінчиться її одиночне скорочення.
Якщо дратівливі імпульси зближені і кожен з них припадає на той момент, коли м'яз тільки почала розслаблятися, але не встигла ще повністю розслабитися, то виникає зубчастий тип скорочення (зубчастий тетанус).
Якщо дратівливі імпульси зближені настільки, що кожен наступний припадає на час, коли м'яз ще не встигла перейти до розслаблення від попереднього подразнення, тобто відбувається на висоті її скорочення, то виникає тривале безперервне скорочення, що одержало назву рівного тетануса.
Гладкий тетанус – нормальний робочий стан скелетних м'язів обумовлюється надходженням з ЦНС нервових імпульсів з частотою 40-50 в 1с.
Зубчастий тетанус виникає при частоті нервових імпульсів до 30 в 1с. Якщо м'яз отримує 10-20 нервових імпульсів в 1с, то вона перебуває в стані м'язового тонусу, тобто помірного ступеня напруги.
Стомлення м'язів. При тривалому ритмічному роздратування в м'язі розвивається стомлення. Ознаками його є зниження амплітуди скорочень, збільшення їх латентних періодів, подовження фази розслаблення і, нарешті, відсутність скорочень при триваючому роздратуванні.
Ще один різновид тривалого скорочення м'язів – контрактура. Вона триває й при знятті подразника. Контрактура м'яза наступає при порушенні обміну речовин або зміні властивостей скорочувальних білків м'язової тканини. Причинами контрактури можуть бути отруєння деякими отрутами і лікарськими засобами, порушення обміну речовин, підвищення температури тіла та інші фактори, що призводять до незворотних змін білків м'язової тканини.
Механізм м'язового скорочення. Вивчення структури м'язових волокон в світловому мікроскопі дозволило виявити їх поперечну смугастість. Електронно-мікроскопічні дослідження показали, що поперечна смугастість зумовлена особливою організацією скорочувальних білків міофібрил – актину, міозину, тропоніна і тропомиозина (рис. 6. У саркомере чергуються поперечні світлі і темні смуги. У центральній частині кожного саркомера розташовані товсті нитки міозину. На обох кінцях саркомера знаходяться тонкі нитки актину. Нитки міозину виглядають в світловому мікроскопі як світла смуга в темному диску, який містить нитки міозину і актину і називається анізотропним або А-диском. По обидві сторони від А-диска знаходяться ділянки, які містять тільки тонкі нитки актину і здаються світлими, вони називаються ізотропними або I-дисками. При скорочення тонкі актінові нитки ковзають уздовж товстих міозинових ниток, рухаючись між ними до середини саркомера; самі актинові і міозинові нитки своєї довжини не змінюють. Міозинові нитки мають поперечні містки з головками, які відходять від ниток біполярно. Актинова нитка складається з двох закручених одна навколо іншої ланцюжків молекул актину. На нитках актину розташовані молекули тропоніна, а в жолобках між двома нитками актину лежать нитки тропоміозина. Молекули тропоміозина в спокої розташовуються так, що запобігають прикріплення поперечних містків міозину до актинових ниток.

Рисунок 6– Організація скелетних м'язів
У багатьох місцях ділянки поверхневої мембрани м'язової клітини поглиблюються у вигляді трубочок всередину волокна, перпендикулярно його поздовжньої осі, утворюючи систему поперечних трубочок (Т-система). Паралельно міофібрилам і перпендикулярно поперечним трубочках розташована система поздовжніх трубочок (альфа-система). Бульбашки на кінцях цих трубочок, в яких зосереджена основна кількість внутрішньоклітинного кальцію, підходять дуже близько до поперечних трубочках, утворюючи з ними так звані тріади. При порушенні ПД поширюється по мембранам Т-системи всередину клітини і викликає вивільнення іонів кальцію з альфа-системи. З появою іонів кальцію в присутності АТФ відбувається зміна просторового положення тропоміна – нитки тропомиозина зсуваються і відкриваються ділянки актину, що приєднують міозинові головки. З'єднання головки міозину з актином призводить до зміни положення містка (його «згинанню»), в результаті нитки актину переміщаються на один мм до середини саркомера. Потім відбувається від'єднання містка від актину. Ритмічні прикріплення та від'єднання головок міозину тягнуть актинову нитку до середини саркомера (рис. 7.
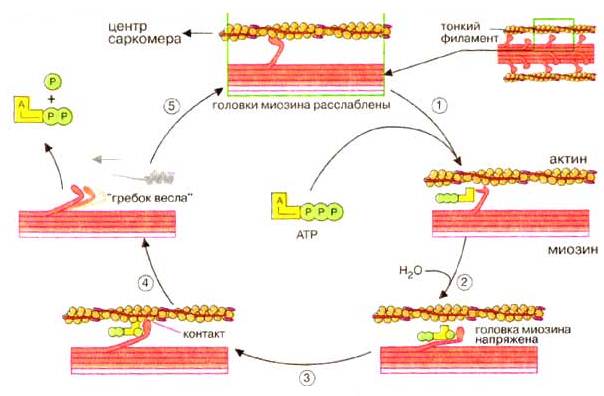
Рисунок 7– Механізм скорочення м'язових волокон
При відсутності повторного порушення іони кальцію закачуються кальцієвим насосом з межфібріллярного простору в систему саркоплазмотичного ретикулума. Це призводить до зниження концентрації іонів кальцію і від'єднання його від тропонина. Внаслідок цього тропомиозин повертається на колишнє місце і блокує активні центри актину. Розслаблення м'яза після її скорочення відбувається пасивно – актинові і міозинові нитки легко ковзають у зворотному напрямку під впливом сил пружності м'язових волокон, а також скорочення м'язів-антагоністів.
Читайте також:
- II. МЕХАНІЗМИ ФІЗІОЛОГІЧНОЇ ДІЇ НА ОРГАНІЗМ ЛЮДИНИ.
- V Процес інтеріоризації забезпечують механізми ідентифікації, відчуження та порівняння.
- V Такі негативні особистісні утворення, як самовпевненість і нерозвиненість автономії та ініціативи, обумовлюють неадаптивне старіння людини.
- А і Б – аферентні волокна.
- Автономні утворення у зарубіжних державах
- Адміністративні зміни кінця 18-19 ст. та утворення нових архівів
- Аеродинамічні властивості колісної машини
- Активний опір ліній електропередачі
- Альдостерон та механізми ренін-ангіотензину
- Аналіз складу майна та джерел його утворення.
- Аналізатори людини та їхні властивості.
- Аналізатори людини та їхні властивості.
| <== попередня сторінка | | | наступна сторінка ==> |
| ВИСНОВОК | | | Будова та функції нервово-м’язового синапсу. |
|
Не знайшли потрібну інформацію? Скористайтесь пошуком google: |
© studopedia.com.ua При використанні або копіюванні матеріалів пряме посилання на сайт обов'язкове. |