- Гідрологія і Гідрометрія
- Господарське право
- Економіка будівництва
- Економіка природокористування
- Економічна теорія
- Земельне право
- Історія України
- Кримінально виконавче право
- Медична радіологія
- Методи аналізу
- Міжнародне приватне право
- Міжнародний маркетинг
- Основи екології
- Предмет Політологія
- Соціальне страхування
- Технічні засоби організації дорожнього руху
- Товарознавство продовольчих товарів
Тлумачний словник
Авто
Автоматизація
Архітектура
Астрономія
Аудит
Біологія
Будівництво
Бухгалтерія
Винахідництво
Виробництво
Військова справа
Генетика
Географія
Геологія
Господарство
Держава
Дім
Екологія
Економетрика
Економіка
Електроніка
Журналістика та ЗМІ
Зв'язок
Іноземні мови
Інформатика
Історія
Комп'ютери
Креслення
Кулінарія
Культура
Лексикологія
Література
Логіка
Маркетинг
Математика
Машинобудування
Медицина
Менеджмент
Метали і Зварювання
Механіка
Мистецтво
Музика
Населення
Освіта
Охорона безпеки життя
Охорона Праці
Педагогіка
Політика
Право
Програмування
Промисловість
Психологія
Радіо
Регилия
Соціологія
Спорт
Стандартизація
Технології
Торгівля
Туризм
Фізика
Фізіологія
Філософія
Фінанси
Хімія
Юриспунденкция
Молекулярні порушення клітини та їх наслідки.
Виділяють три групи порушень молекул у клітині: 1) втрата здатності молекулярних структур виконувати властиві їм у нормі функції; 2) втрата здатності відповідних молекул забезпечувати структурні перебудови хромосом, що супроводжують поділ клітини; 3) зміна кодового, інформаційного та функціонального значення послідовності мономерів у біологічних макромолекулах.
Із зазначеними ушкодженнями макромолекул асоціюються такі прояви реакцій клітини на опромінення, як відхилення від норми метаболічних процесів, генетичної регуляції функцій клітини й генетичні порушення, що можуть проявлятися в наступних поколіннях, у нащадків опроміненого організму.
Ушкодження молекули ДНК проявляються в різних молекулярно-біологічних і клітинних реакціях, які є мірою радіобіологічного ефекту. Розриви ДНК. Розрізняють:
одинарні розриви, коли зв'язок між окремими атомними угрупуваннями порушується в одній із ниток двониткової молекули ДНК. Виникнення їх пропорційне дозі опромінення. Поодинокі розриви, як правило, не призводять до пошкодження молекули ДНК, оскільки розірвана нитка утримується на місці водневими, гідрофобними та іншими видами взаємодії з протилежною ниткою ДНК. Крім того, ці розриви досить легко усуваються системами відновлення клітини − так званими системами репарації.
подвійні розриви – з підвищенням дози опромінювання зростає ймовірність одинарних розривів у близьких ділянках обох ниток ДНК і формується подвійний розрив, що призводить до розпаду молекули ДНК. Виникнення подвійних розривів пропорційне квадрату дози опромінення. У разі опромінення клітин щільноіонізуючими випромінюваннями (альфа-частки) спостерігається прямо пропорційна залежність появи подвійних розривів від дози опромінення, оскільки відбувається пошкодження обох ниток ДНК однією зарядженою часткою. Подвійні розриви призводять до аберацій хромосом, що є причиною летального ефекту опромінення клітин.
Формування хромосомних аберацій. Найвиразніше проявляються ушкодження структури хромосомного апарату клітин після їх опромінення. Структура хромосом змінюється внаслідок того, що в разі спотворення молекулярної структури ДНК або хроматину внеможливлюється звичайний хід реплікації, формування хромосом і здійснення всіх процесів мітотичного поділу клітини.
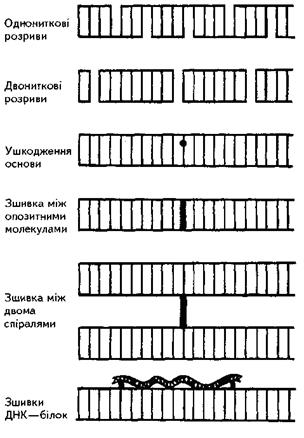
Рис. 4. Основні типи молекулярних ушкоджень ДНК
Зміни структури хромосомного апарату називають хромосомними абераціями (перебудовами, або мутаціями). Вони, (рис. 5) можуть виражатись у зміні числа генів у хромосомах унаслідок або випадіння з хромосоми її частини − делеції, або подвоєння ділянок хромосом − дуплікації. Якщо ж в одній із ділянок хромосоми гени розташовані у зворотному порядку порівняно з нормою, то це явище називають інверсією. В структурі ділянки хромосоми, що зазнала інверсії, може бути центромера, й тоді йдеться про парацентричну інверсію. Коли інвертована ділянка не має центромери, інверсію називають перицентричною.
До хромосомних аберацій належить також зміна положення ділянки хромосоми в хромосомному наборі. Аберації цього класу називають транслокаціями. Найпоширенішими є реципрокні транслокації, за яких відбувається обмін ділянками між негомо-логічними хромосомами. Якщо ділянка хромосоми змінює своє положення без реципрокного обміну й залишається в тій самій хромосомі або ж потрапляє в іншу, то йдеться про транспозицію.
Транслокація може бути збалансованою, коли до розривів хромосоми приєднуються однакові фрагменти іншої хромосоми. При цьому виникають так звані робертсонові структури, серед яких трапляються й дицентричні хромосоми. Утворення хромосомних аберацій цього типу активізується під впливом опромінення. Поява робертсонових структур не спричиняє захворювання. Натомість у разі незбалансованої транслокації спостерігається фізична або ментальна недостатність.
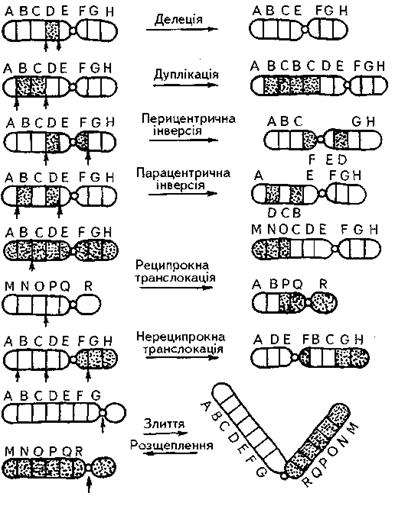
Рис.5. Основні типи перебудов хромосом
Типи хромосомних перебудов залежать від фази клітинного циклу. Якщо розриви хромосоми виникли у фазі G1, коли вона представлена однією ефективною ниткою, утворюються хромосомні аберації:
- хромосомні симетричні і асиметричні транслокації(структурні зміни хромосом, коли хромосомний сегмент - локус переноситься на інше місце);
- кінцеві делеції (втрата ділянки хромосом);
- парацентричні і перицентричні інверсії, якщо розриви виникають після синтезу ДНК, коли хромосома складається з двох хроматид (фази S і G2), виникають хроматидні аберації: ізолокусні розриви, складні хроматидні симетричні і асиметричні транслокації, дуплікації (подвоєння тієї самої хромосоми). Лише симетричні транслокації й інверсії можуть передаватися в ряді поколінь клітин. Решта спектра аберацій виявляється в першому після опромінення мітозі у вигляді мостів, фрагментів, кільцевих хромосом, що призводять до втрати генетичного матеріалу, внаслідок чого настає репродуктивна інактивація клітин.
Підрахунок кількості клітин з абераціями хромосом використовують як кількісний метод оцінки летального ушкодження клітин.
У разі опромінення клітини в пресинтетичній фазі фрагментації й обміни фрагментами як процеси, пов'язані з розривами молекули ДНК, стосуються всієї хромосоми − носія однієї молекули ДНК. Аберації, які виникають при цьому, називають хромосомними (рис.6). Якщо ж фрагментації й обміни індукуються в реплікативній або пост-реплікативній фазі, то внаслідок індивідуальності молекул ДНК у хроматидах аберації хромосом спричиняються розривами молекули ДНК у хроматидах. Такі аберації називають хроматидними.

Рис.6. Види аберацій хроматичного типу в різних фазах мітозу
Механізми виникнення хромосомних аберацій.Намагання пояснити природу процесів, які завершуються формуванням хромосомних аберацій, зумовили створення кількох гіпотез.
Класична гіпотеза, появі якої сприяли дослідження відомого генетика Л. Дж. Стадлера, проведені ще в 20-х роках XX ст., стверджувала, що після впливу іонізуючого випромінювання на хромосоми виникають фрагменти з «липкими», «незагоєними» кінцями, котрі з'єднуючися в різних комбінаціях, дають усе різноманіття хромосомних аберацій.
Пізніше було введено поняття потенційних ушкоджень хромосом, тобто порушень, що можуть із часом «заліковуватися» й не перетворюватися на розриви. Приводом для введення цього поняття послужили численні експериментальні факти, що свідчили про можливість впливу на вихід хромосомних аберацій різних за природою дії факторів. Зазначене поняття покладено в основу гіпотези потенційних ушкоджень.
Із потенційними ушкодженнями хромосом пов'язують передмутаційні зміни клітин. Отже, згідно з гіпотезою потенційних ушкоджень, є принаймні три етапи формування хромосомних аберацій: 1) поява в клітині потенційних ушкоджень; 2) часткове відновлення клітини від потенційних ушкоджень; 3) перетворення потенційних ушкоджень на невідновлювані.
Були спроби класифікувати потенційні ушкодження за тривалістю їх існування. Дослідження молекулярно-біологічних механізмів дії іонізуючої радіації на ДНК показали, що ушкодженнями, реалізація яких призводить до формування хромосомної аберації, є подвійні розриви ДНК. Тому часто розриви хромосом ототожнюють із подвійними розривами молекули ДНК. Проте слід ураховувати, що розрив хромосоми, який, безперечно, пов'язаний із двонитковими розривами ДНК, охоплює також перетворення четвертинних структур хроматину.
Гіпотеза помилкової репарації пов'язує формування хромосомних аберацій із наслідками хибних дій ферментів репарації, які в разі появи однониткового розриву ДНК можуть помилково надрізати й другу нитку в опозитному положенні, що призводить до двониткового розриву. Появу потенційних ушкоджень автори цієї гіпотези вбачали у виникненні пробілів.
Дуже поширеною є обмінна гіпотеза Рівелла, згідно з якою аберації хромосом виникають на основі механізму, подібного до кросинговеру. Ушкодження ДНК унаслідок опромінення спонукають хромосоми до неспецифічної асоціації, яка супроводжується обмінами фрагментами. Гіпотеза Рівелла неодноразово змінювалася й доповнювалася, бо експериментальні дані далеко не завжди відповідали теоретичним передбаченням, що випливали з обмінної гіпотези. Проте доповнення до цієї гіпотези не нівелювали основної ідеї про подібність процесу формування хромосомних аберацій до кросинговеру.
Як бачимо, пояснення механізмів виникнення хромосомних аберацій поки що є гіпотетичними. Разом із тим виходи хромосомних аберацій різних типів дуже детально досліджено на різних видах тварин і рослин. Очевидно, вихід хромосомних аберацій є чи не найчастіше застосовуваним параметром для кількісної оцінки радіобіологічних ефектів.
Формування хромосомних аберацій здебільшого внеможливлює подальший поділ клітини, яка внаслідок цього зазнає проліферативної загибелі, й із часом після опромінення в клітинній популяції не буває потомків клітин із абераціями. Виняток становить особливий тип аберації − клітини з дицентричними хромосомами, тобто хромосомами з двома центромерами, за якої клітини зберігають здатність до поділу.
Появу дицентричних хромосом, що тривалий час зберігаються в клітинних лініях від опромінених клітин, використовують як маркери радіаційного впливу. Ці хромосоми в ході мітотичного поділу клітини досить часто утворюють мости, які можуть розриватися, внаслідок чого формуються ядра, що мають тонкий виріст у простір цитоплазми. Такі ядра назвали «хвостатими». Частота їх виявлення є чітко дозозалежною й корелює з частотою хромосомних аберацій у лімфоцитах опромінених тварин і людини.
Часові характеристики формування хромосомних аберацій.Тривалість формування хромосомних аберацій залежить від фази, в якій перебуває клітина в момент опромінення, й типу аберації. Наприклад, у клітин традесканції час формування аберацій варіює від 5 до 30хв, в інших видів − становить близько 1,5 год, а в клітин тварин − 1 год, хоча буває, що процес триває до 4 год.
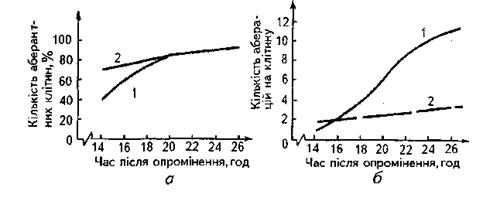
Рис.7. Час появи аберантних клітин (а) і хромосомних аберацій (б) після опромінення клітин китайського хом’ячка у фазі G1
1 – прискореними ядрами криптону 1 х 106 см -2; 2 – ренгенівським випромінюванням (7Гр).
Під впливом іонізуючого випромінювання зростає частота появи мультиаберантних клітин, тобто клітин, в яких сформувалося кілька хромосомних аберацій (рис. 9.8). Збільшення числа аберацій в одній аберантній клітині є наслідком поступового нагромадження мутаційних змін, яке відбувається не тільки під час опромінення, а й триває після нього.
Теломери у формуванні хромосомних аберацій.У нормі на фізичних кінцях хромосом є теломери — структури, що складаються з білків і коротких нуклеотидних послідовностей (ТТААПТ)n, що тандемно повторюються.
Фізіологічна функція теломери полягає в тому, щоб запобігти злиттю або позаплановій рекомбінації хромосом. У разі розривів хромосоми втрачають теломери й набувають нестабільності, яка реалізується в рекомбшаційних процесах і злитті хромосом. Хромосоми можуть відновлювати теломери, синтезуючи їх по розривах. Але за відсутності теломер відбувається й злиття хромосом, і обмін їхніми фрагментами. Зокрема, подвійні хромосомні розриви супроводжуються транслокацією, обмінами фрагментами між двома хромосомами.
4. Механізми формування геномних та точкових мутацій.
Переглядів: 2547