- Гідрологія і Гідрометрія
- Господарське право
- Економіка будівництва
- Економіка природокористування
- Економічна теорія
- Земельне право
- Історія України
- Кримінально виконавче право
- Медична радіологія
- Методи аналізу
- Міжнародне приватне право
- Міжнародний маркетинг
- Основи екології
- Предмет Політологія
- Соціальне страхування
- Технічні засоби організації дорожнього руху
- Товарознавство продовольчих товарів
Тлумачний словник
Авто
Автоматизація
Архітектура
Астрономія
Аудит
Біологія
Будівництво
Бухгалтерія
Винахідництво
Виробництво
Військова справа
Генетика
Географія
Геологія
Господарство
Держава
Дім
Екологія
Економетрика
Економіка
Електроніка
Журналістика та ЗМІ
Зв'язок
Іноземні мови
Інформатика
Історія
Комп'ютери
Креслення
Кулінарія
Культура
Лексикологія
Література
Логіка
Маркетинг
Математика
Машинобудування
Медицина
Менеджмент
Метали і Зварювання
Механіка
Мистецтво
Музика
Населення
Освіта
Охорона безпеки життя
Охорона Праці
Педагогіка
Політика
Право
Програмування
Промисловість
Психологія
Радіо
Регилия
Соціологія
Спорт
Стандартизація
Технології
Торгівля
Туризм
Фізика
Фізіологія
Філософія
Фінанси
Хімія
Юриспунденкция
Компоненти ферментного комплексу
Схема мейоза
Ядерце
Nm
Н1 nDNA
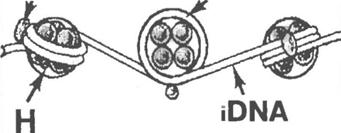
Ділянки ДНК, "намотані" на гістонові октамери, мають довжину в 140 нуклеотидних пар (н. п.) і називаються коровими (core -ДНК, або nDNA).
Нуклеосома розташована на молекулі ДНК не впритул: між ними є лінкерні (сполучні) ділянки (iDNA) завдовжки 60 нуклеотидних пар. З кожною лінкерною ділянкою пов'язана 1молекула ще одного виду гистонів – H1.
Отже, усього в хромосомах зустрічаються гістони 5 видів, а період нуклеосомної організації складає 200 нуклеотидних пар.
Молекула ДНК бере участь в утворенні дуже великої кількості нуклеосом (в середньому 600 000). В результаті на даному рівні організації кожна хромосома (не рахуючи негістонових білків) є довгою ниткою "намистинок" - нуклеосом діаметром 10 нм. В порівнянні з молекулою ДНК, довжина нуклеосомної нитки приблизно в 6,2 рази менше.
У інтерфазному ядрі хромосоми не помітні, а сприймаються всі разом як хроматин. При цьому виділяють гетеро-іеухроматин.
Гетерохроматин - сильно конденсовані і тому функціонально неактивні ділянки хромосом. Вони мають вигляд щільних глибок (кульок) і інтенсивно фарбуються базофільними барвниками. Більшість глибок знаходяться на периферії ядра і ближче (майже впритул) до ядерної оболонки.
Навпаки, еухроматин - функціонально активні, практично деконденсовані і тому світлі ділянки хромосом, розташовані між глибками гетерохроматина.
Нуклеосомний рівень організації є, мабуть, і в гетеро-, і в еухроматині. Але в тих локусах еухроматину, на яких в даний момент часу функціонують ферментні комплекси (реплікації, репарації або транскрипції), як вважають, ДНК вивільняється з взаємодії з гістонами. Тобто тут нуклеосомная організація тимчасово зникає, щоб згодом знову відновитися.
На відміну від цього, в гетерохроматині до нуклеосомному рівня додаються наступні рівні укладання хромосоми. Вважається, що нуклеосомная нитка закручується в спіраль типу соленоїда, а та, можливо, утворює суперспіраль. У цих процесах, мабуть, ключову роль грає гістон H1. У результаті формується хроматинова нитка діаметром 30 нм.
Хроматинова нитка коротша нуклеосомної приблизно в 18 разів і коротша упакованої в ній молекули ДНК в 6,2 х 18 = 100 разів.
У свою чергу, хроматинові нитки утворюють петлі, які збираються в розетки, де основи петель кріпляться до білків ядерного матриксу. У гетерохроматині такі групи петель більш менш щільно прилягають один до одного.
Найбільшої компактизації хромосоми досягають в процесі мітоза (на стадії метафази). Точне укладання хромосомних ниток при цьому невідома.
За однією з версій, хроматинова нитка багаторазово складається по довжині хромосоми (мал. 1.4).

Тому при мікроскопії на поперечному зрізі виявляється близько 100 хроматинових ниток (що є перерізами однієї і тієї ж нитки).
Крім того, петлі хроматинової нитки мають довжину не усієї хромосоми, а лише окремих її сегментів - хромомерів. Це пояснює можливість сегментації хромосом при тих або інших діях.
Метафазні хромосоми
Морфологію хромосом зазвичай характеризують по їх стану на стадії метафази мітоза (мал. 1.5).
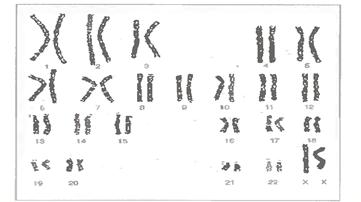
Загальна довжина усіх 46 метафазних хромосом приблизно в 100 000 разів менша загальної довжини (- 190 см) молекул ДНК, які містяться в них, і складає близько 180 мкм.
Це слідує і з наведеного вище опису: перехід від ДНК до хроматинової нитки, так і згортання останнього в метафазную хромосому дає 100-кратне скорочення довжини.
Майже у кожної хромосоми виявляються наступні частини:
а) центромера(первинна перетяжка),
б) плечі- частини хромосоми по сторонах від центромери,
в) теломери - кінцеві ділянки плечей.
У області центромери знаходиться кінетохор - місце прикріплення клітинного веретена.
По положенню центромери хромосоми діляться на 3 види:
а) метацентричні - з рівними плечима,
б) субметацентричні- з плечима неоднакової довжини,
в) акроцентричні- одне плече практично відсутнє.
В цілому ж за розміром і формою хромосоми людини поділяються на 7 груп, показаних на рисунку.
Негістонові білки хромосом
Загальний масовий вміст кислих (негістонових) білків в хромосомах істотно менший, ніж гістонів. Проте ці білки надзвичайно різноманітні (включають, принаймні, декілька сотень різних представників).
Ймовірно, деякі з кислих білків відіграють структурну роль, беручи участь в утворенні наднуклеосомних рівнів укладання хромосом.
Іншу групу складають численні ферменти, що забезпечують процеси реплікації, модифікації, репарації і транскрипції.
Нарешті, найрізноманітнішою по складу, мабуть, є група регуляторних білків. Вони контролюють активність вищезгаданих ферментів, а також доступність тих або інших ділянок ДНК для цих ферментів.
З конкретними представниками двох останніх груп негістонових білків хромосом ми неодноразово будемо стикатись.
Ядерце (нуклеола) є найщільнішою структурою ядра і зазвичай має округлу форму. У ядрі може міститься як одне, так і декілька ядерець.
В той же час ядерце - це не окрема від хроматину структура, а його похідна. Воно формується у зв'язку з певними ділянками хромосом – так званими організаторами ядерця.
Кожен такий організатор містить декілька десятків або сотень копій генів рибосомної РНК (рРНК). На цих генах активно відбувається синтез попередників рРНК (про-РНК).
Останні тут же (у ядрі) піддаються дозріванню і, зв’язуються з рибосомальними білками, утворюють субодиниці рибосом.
Відповідно до вищесказаного, при електронній мікроскопії (мал. 1.6) біля ядерця виявляється пов'язана з ним ділянка хроматину (Сhr). У самому ж ядерці виявляються фібрилярні компоненти (про-рРНК, рРНК і їх гени на ДНК), а також гранулярні структури (субодиниці рибосом).
А при світловій мікроскопії ядерця відрізняються базофілією (із-за великої концентрації РНК) і позитивним забарвленням по Браше (на РНК).
Від ядерця субодиниці рибосом переміщуються до пор ядерної оболонки, через які проходять в цитоплазму.

1.2. РЕПЛІКАЦІЯ ОСНОВНОЇ ЧАСТИНИ ДНК
Один з найважливіших внутрішньоядерних процесів - реплікація, або подвоєння кількості ДНК. Часто цей процес називают також реплікативним синтезом ДНК (маючи на увазі, що буває ще репаративний синтез).
Саме реплікація ДНК забезпечує відтворення спадкової інформації при утворенні нових клітин.
Приблизно в той же час відбувається подвоєння кількості і хромосомних білків. Отже, реплікація ДНК - складова частина більш складнішого процесу - реплікації хромосом.
Місце реплікації ДНК в клітинному циклі
Схеми мітоза і мейозу
Реплікативний синтез ДНК тісно пов'язаний з поділом клітини.
Як відомо, клітинні поділи бувають двох типів: мейоз і мітоз.
Мейоз використовується лише в одному випадку: так проходит останній поділ попередників статевих клітин. Інші поділи здійснюються шляхом мітоза; це всі попередні поділи статевих клітин, а також усі поділи соматичних клітин.
Якщо позначити кількість ДНК в одинарному наборі хромосом через n, то його зміна в ході вказаних двох типів поділі відіб'ється в хромосомному наборі.
| |||
| |||
Схема мітоза
У обох випадках ядро початкової клітини містить звичайний подвійний набір хромосом і в ньому - диплоїдна (2n) кількість ДНК. При підготовці до поділу ця кількість шляхом реплікації зростає до тетраплоїдної (4n), а в ході наступного поділу розподіляється порівну між двома дочірніми клітинами.
Але при мейозі додається ще один поділ, причому без попередньої реплікації ДНК; отже у результаті утворюються чотири клітини з одинарним, або гаплоїдним (n), числом ДНК.
Отже, серед усіх поділів еукаріотичних клітин лише в одному випадку (у другому поділі мейозу) не виникає попереднього подвоєння ДНК.
Відмінним є і той період клітинного циклу, коли здійснюється реплікація ДНК.
У мейотичних клітинах (сперматоцитах і овоцитах) синтез ДНК вважають початком профази першого поділу, а саме - прелептотенною стадією профази. Потім слідує лептотенна стадія профази (конденсація хромосом). Причому в цілому профаза мейозу (що включає шість стадій) триває дуже довго - близько місяця в сперматоцитах і до декількох десятиліть (включаючи тривалий період спокою) в овоцитах.
У мітотичних же клітинах, навпаки, між реплікацією ДНК і початком профази розрізняють певний проміжок часу, а сама профаза проходить досить швидко (не більше декількох годин).
Мітотичний цикл
У результаті життєвий цикл клітини (мітотичний цикл), що мітотично діляться, поділяють наступним чином (мал. 1.8).
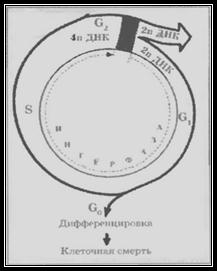
S-період: період, коли в ядрі відбувається реплікація практично усієї ДНК. Нереплікованими, напевно, залишаються лише центромерні ділянки ДНК: вони подвоюються на початку анафази мітозу.
У сперматогоніях S-період триває близько 15 годин; у інших клітинах він може тривати менше (8 годин) або більше. Для порівняння: в мейотичних клітинах сперматоцитах - подвоєння ДНК займає близько 100 годин.
Упродовж S-періоду кількість ДНК в ядрі поступово зростає від 2n до 4n.
У S-період подвоюється також кількість хромосомних білків в ядрі, а біля ядра відбувається дуплікація центріолей.
G2-період - постсинтетичний, або премітотичний. Зазвичай він не дуже тривалий і включає синтез ряду інших речовин, необхідних для проходження мітозу. В цей час активно синтезується білок мікротрубочок тубулін, який використовується для формування веретена поділу.
Вміст ДНК в цей період – 4n.
Потім відбувається мітотичний поділ (на схемі - М), що включає чотири фази (профазу, метафазу, анафазу, телофа-зу), що призводить до утворення двох диплоїдних клітин.
Нарешті,G2-період - постсинтетичний, або премітотичний. Це деякий інтервал часу від закінчення мітоза до початку синтезу ДНК (і ядерних білків) в дочірній клітині. У цей період відновлюється вміст цитоплазматичних білків і, як наслідок, відбувається ріст клітини (до разміру материнської). Вміст ДНК в клітині – 2n.
Стадії , G1, S, G2 разом складають інтерфазу.
Типи клітин по здатності до поділу.
Перераховані стадії життєвого циклу відносяться лише до клітин, що регулярно діляться.
Взагалі ж по здатності до поділів усі клітини дорослого організму (окрім спермато- або овоцитів) поділяються на 3 типи.
а) Мітотичні, тобто клітини, що постійно діляться, :
- більшість клітин базального шару епітелію
- гемопоетичні клітини початкових стадій дозрівання
- сперматогонії і деякі інші клітини.
б) Умовно постмітотичні клітини. Це клітини, які не діляться, що зберегли здатність до поділів при дії певних стимулів. Частіше усього поділи поновлюються при регенерації відповідного органу або тканини.
Сюди відносяться клітини печінки, а також стовбурові клітини кісткових, скелетних, м'язових і деяких інших тканин.
в) Постмітотичніклітини - клітини, що не діляться, остаточно втратили здатність ділитися.
Приклади - клітини усіх шарів епідермісу шкіри, окрім базального; нервові клітини, клітини серцевого м'яза, симпласти (волокна) скелетних м'язів.
Деякі цитологи вважають за краще поділяти не клітини по здатності до поділів, а популяції клітин.
Зокрема, клітини мозку і симпласти (волокна) скелетних м'язів позначаються як стаціонарні популяції.
Клітини усіх шарів епідермісу (включаючи базальний), гемопоетичні і тому подібне - називаються популяціями, що оновлюються.
А клітини залозистих органів (наприклад, печінки) розглядають як популяції, які ростуть .
У подібній термінології є принциповий недолік: вона абсолютно спотворює суть справи.
Наприклад, саме так звані оновлювані популяції знаходятся в стаціонарному стані.
Дійсно, стаціонарним є такий динамічний стан, при якому за рахунок рівності в системі "Притоку" і "відтоку" вміст кожного компонента залишається на постійному рівні. І популяція гемопоетичних клітин, і популяція епідермальних клітин - типові представники стаціонарних систем.
Навпаки, ті популяції, які названі стаціонарними, якраз нестаціонарні. Наприклад, в мозку чисельність нейронів, хоча і повільно, але монотонно знижується. Отже швидше підійшов би термін "квазістаціонарні" або "Статичні" популяції.
Нарешті, так звана зростаюча популяція клітин печінки в звичайних умовах зовсім не росте.
Вихід клітин з мітотичного циклу
Про клітини, що виходять з мітотичного циклу, говорять, що вони вступили в G2-період свого життєвого циклу. Якщо не рахувати "сплячих" стовбурових клітин, такий вихід, зазвичай, призводить в результаті диференціювання. Це можна представити наступними схемами:
М→ G1 ↔G0 ↔ G0(D1) → G0(D2) →G0 (D3);
М →G1 ↔ G0 ↔G0(D1) →G0 (D2) →G0 (D3) → F → загибель.
Тут G0(D1)- різні стадії диференціювання; причому, на усіх цих стадіях клітини не діляться (знаходяться в G0-періоді). Проте клітини G0 і G0(D1) є умовно постмітотичними, тобто здатні повернутися в мітотичний цикл і реплікувати ДНК. У клітин же наступних стадій диференціації така здатність втрачена.
У першому прикладі зрілі клітини функціонують в організмі велику частину його життя (нервові клітини, кардіоміоцити).
У другому прикладі має місце термінальна диференціація, що закінчується загибеллю клітин і зміною їх іншими геніраціями таких же клітин (кератиноцити, клітини крові).
З вищевикладеного виходить, що реплікативний синтез ДНК відбувається в S-фазі циклу мітотичних клітин, а при деяких обставинах здійснюється в умовно постмітотичних клітинах. Крім того, в список синтезуючої ДНК клітин потрібно додати сперматоцити і овоцити на стадії прелептотени.
Загальна характеристика реплікації ДНК
Основні принципи
Реплікація ДНК має ряд принципових особливостей.
а) По-перше, субстратами, з яких синтезуються нові ланцюги ДНК, є дезоксинуклеозидтрифосфати (дНТФ), а не дезоксинуклеозидмонофосфати (дНМФ), які входять до складу ДНК.
Тому в ході включення в ланцюг ДНК від кожного нуклеотиду відщепляються 2 фосфатні залишки (у вигляді пірофосфату, який незабаром гідролізується до фосфатів) :
вільні дНМФ →залишки дНТФ в нових ланцюгах ДНК + пірофосфат
Використання саме дНТФ, а не дНМФ, пояснюється енергетичними причинами: утворення міжнуклеотидного зв'язку вимагає енергії; джерелом її і служить розрив міжфосфатного зв'язку.
б) По-друге, реплікація ДНК - матричний процес: кожен ланцюг ДНК, що синтезується (дочірня), будується, використовується як матриця один з ланцюгів початкової (батьківської) ДНК.
Основою при цьому є принцип комплементу: з чотирьох можливих нуклеотидів (дАТФ, дГТФ, дЦТФ, дТТФ) до складу зростаючого ланцюга включається в даний момент той, який комплементарний нуклеотиду у відповідному положенні і батьківському ланцюгу.
в) В третіх, процес (на відміну, наприклад від синтезу РНК) є симетричним: матрицями служать обидва ланцюги батьківської ДНК (мал. 1.9).
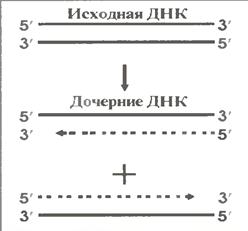
Також його можна назвати напівконсервативним: після закінчення процесу вихідні молекули ДНК будуть наполовину оновленними. У кожній з дочірних молекул один ланцюг - батьківський (на мал. 1.9 показана суцільною лінією), а друга – новосинтезована (пунктирна лінія).
г) Нарешті, дуже важливий момент торкається напрямку зростання і полярності ланцюгів ДНК.
Подовження ланцюга ДНК (чи окремого її фрагмента) завжди відбувається в напрямі від 5'- кінця до 3'- кінця. Це означає, що черговий новий нуклеотид приєднується до 3'- кінця зростаючого ланцюга.
Крім того, оскільки в будь-якій молекулі ДНК комплементарні ланцюги антипараллельні, то і зростаючий ланцюг антипараллельна матричному ланцюгуа. Отже, остання прочитується у напрямі 3' → 5'.
Особливості механізму
Відмітимо ще декілька менш принципових, але достатньо важливих особливостей, які можна віднести до механизму реплікації ДНК.
а) Процес реплікації здійснюється складним ферментним комплексом (що налічує до 15-20 різних білків). Ключові компоненти цього комплексу ми вкажемо пізніше. Зараз же підкреслимо, що при реплікації ДНК у еукаріот на кожній хромосомі працює не один, а відразу велика кількість таких комплексів.
Іншими словами, на хромосомі є багато точок початку реплікації ДНК. І подвоєння ДНК здійснюється не послідовно від одного кінця до іншого, а одночасно в багатьох місцях відразу.
Це значно скорочує тривалість процесу. Так, в сперматогоніях на одній хромосомі в середньому близько 40 точок початку реплікації, і S-фаза триває, як вже відзначалося, 15 годин. На відміну від цього, в прелептотенних сперматоцитах хромосоми мають в середньому всього по 5-6 таких точок, тому реплікація подовжується до 100 годин.
б) В кожній вказаній точці починають працювати два ферментних комплекси: один переміщається по молекулі ДНК в одну сторону, другий - в протилежну.
При цьому кожен комплекс реплікує не лише один ланцюг ДНК, але і іншу. Найскладніше питання: як при цьому вдається для обох батьківських ланцюгів (незважаючи на їх антипаралельність) дотримувати принцип комплементарності у напрямі 3' →5'. Можливі механізми ми коротко обговоримо нижче.
Але який би не був механізм, реплікація поширюється в обидві сторони від кожної точки початку реплікації (мал. 1.10).
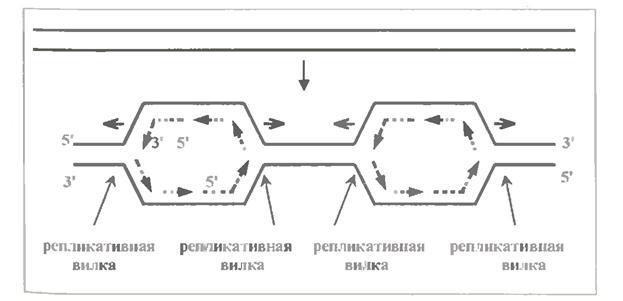
Говорять, що при цьому утворюються дві репликативні вилки, які рухаються в протилежних напрямах. Між цими вилками з'являється "вздуття", що поступово розширюється, або "очко" : це вже репліковані відділи ДНК.
Кінець кінцем сусідні зони реплікації ("вздуття") зливаються і уся молекула ДНК виявляється подвоєною.
в) Ферментний комплекс функціонує так, що один з двох синтезуємих ним ланцюгів, росте з деяким випередженням в порівнянні з іншим ланцюгом. Відповідно, перший ланцюг називается лідируючим, а другий – запізнюючим.
Найважливіше полягає в тому, що лідируючий ланцюг утворюється ферментним комплексом у вигляді безперервного дуже довгого фрагмента. Його довжина (у нуклеотидах), очевидно, дорівнює половині відстані між двома сусідніми точками початку реплікації. Для сперматогоній це близько 1 600 000 нуклеотидів. На мал. 1.10 такі фрагменти показані довгими переривчастими стрілками.
Запізнюючий ланцюг утворюється у вигляді серії відносно коротких фрагментів - приблизно по 1500 нуклеотидів. Це так звані фрагменти Оказаки (на малюнку зображені короткими переривчастими стрілками).
З мал. 1.10 видно: у вигляді фрагментів Оказаки синтезується ферментним комплексом той ланцюг, напрямок утворення якого протилежний напряму руху відповідної реплікативної вилки.
Так, крайня ліва на малюнку вилка переміщується теж вліво. Для верхньої із зростаючих ланцюгів це співпадає з напрямом її росту : 5' → 3'. Тому цей ланцюг є таким, що лідирує і росте у вигляді довгого безперервного фрагмента.
А для нижньої із зростаючих ланцюгів те ж, єдино разрешенное, напрям зростання (5´ → 3') протилежний до напряму руху лівої вилки. Відповідно, цей ланцюг - запізнюється і формується у вигляді коротких фрагментів Оказаки. Очевидно, таким чином ферментній системі легше здолати утруднення, пов'язані з неспівпаданням вказаних напрямів.
Відмітимо, що у разі сусідньої реплікативної вилки положенння лідируючого і запізнюючого ланцюгів обернено попередньому. Тут вже нижній ланцюг є лідируючим, а верхній - запізнюючим і представлений фрагментами Оказаки.
г) Нарешті, остання в цій групі подія.
Утворенню кожного фрагмента ДНК (як довгого, так і будь-якого з фрагментів Оказаки) передує синтез короткої послідовності (з 10-15 нуклеотидів) РНК-приманки (РНК-затравки).
Річ у тому, що основний фермент, що синтезує ДНК (ДНК-полімераза), не може починати процес "з нуля", тобто у відсутністі олігонуклеотидної послідовності. На противагу цьому, фермент синтезу РНК (РНК-полімераза) таку здатність має. Через те цьому ферменту і "доводиться" починати утворення кожного нового фрагмента ДНК.
Для синтезу РНК-приманки необхідні рибонуклеозидтрифосфати (рНТФ), а їх включення відбувається теж за принципом комплементу відповідній ділянці ДНК.
РНК-послідовності відрізняються від ДНК-послідовностей лише двома відмінностями: в нуклеотидах пентоза містить в положенні 2´ гідроксильну групу, а в четвірці азотистих основ тимін замінений на урацил (позбавлений, в порівнянні з тиміном, метильної групи).
Але ці дві відмінності істотно позначаються на здатності утворювати дволанцюгову структуру. Тому послідовність РНК-приманки після завершення синтезу фрагментів ДНК видаляється. Замість них відбувається добудовування (шляхом подовження попереднього фрагмента ДНК) "проломів", що утворилися.
І, нарешті, усі численні фрагменти ДНК, утворені на одному батьківському ланцюзі, зшиваються в єдиний ланцюг.
Як вже відзначалося, в процесі реплікації ДНК бере участь складний ферментний комплекс, що включає, по деяким оцінкам, 15-20 білків.
Але функція і механізм дії доки виявлені не для усіх цих білків, тому в нижченаведеному описі фігурує "лише" 12 найменувань.
Для зручності викладу розділимо перераховувані білки на 3 групи (мал. 1.11).
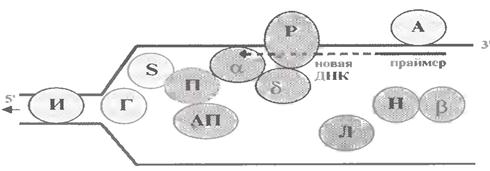
Білки, що готують батьківську ДНК до реплікації
а) Точки початку реплікації на молекулі ДНК мають специфичну послідовність основ, багату парами А-Т.
Процес починається з того, що з кожною такою послідовністю зв'язується декілька молекул спеціальних впізнавальних білків. У бактерій такі білки називаються DnaA (як перші білки, що ініціюють реплікацію). Тому на мал. 1.11 білок, що впізнає, позначений буквою А.
Можна уявити різні причини, по яких можлива взаємодія білків, що впізнають, з точками початку реплікації. Серед цих причин:
- сама поява в ядрі білків, що впізнають, або їх певна модифікація;
- звільнення точок початку реплікації від деяких блокуючих елементів;
- поява в ядрі якихось третіх чинників, необхідних для даної взаємодії; і т. д.
Наявні дані свідчать на користь першого варіанта. Але у будь-якому випадку ясно, що тут - одна з ключових ланок, контролюючих початок реплікації.
Білки, що впізнають, що забезпечують зв'язування ДНК-реплікуючого комплексу, мабуть, далі не переміщуються разом з ним по ДНК.
б) Одним з "першопроходців" виступає фермент геліказа (від helix - спіраль; на мал. 1.11 позначений буквою Г). Він забезпечує розплітання в районі реплікативної вилки подвійної спіралі батьківської ДНК : остання відокремлюється на одноланцюгові ділянки.
На це витрачається енергія гідролізу АТФ - по 2 молекули АТФ на розподіл 1 пари нуклеотидів.
Мабуть, одночасно відбувається також витіснення даної ділянки ДНК із зв'язку з гістонами і іншими хромосомними білками.
в) Проте розплітання спіралі на деякій ділянці створює суперспіралізацію перед цією ділянкою.
Річ у тому, що кожна молекула ДНК в цілому ряді місць зафіксована на ядерному матриксі. Тому вона не може вільно обертатися при розплітанні якоїсь своєї ділянки. Це і викликає суперспіралізацію, а з нею - утворення структурної напруги, блокуючої подальше розплітання подвійної спіралі.
Проблема вирішується за допомогою ферментівтопоізомераз (І на мал. 1.11). Очевидно, вони функціонують на ще нерозплетеній ділянці ДНК, тобто там, де виникає суперспіралізація.
Так звана топоізомераза I розриває один з ланцюгів ДНК, переносить її проксимальний кінець на себе (мал. 1.12).
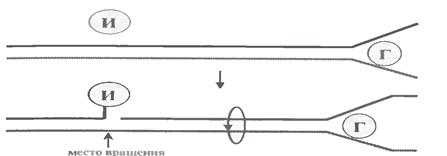
Це дозволяє дистальній ділянці ДНК (від місця розплетення до місця розрива) обертатися навколо відповідного зв'язку цілого ланцюга, що і попереджує утворення супервитків. Згодом кінці розірваного ланцюга знову замикаються: один з них переноситься з ферменту на другий кінець. Отже процес розриву ланцюга топоізомеразою зворотній.
Є також топоізомераза II (бактеріальна топоізомераза II називається гіразою). Цей фермент розриває відразу обидва ланцюги ДНК, знову-таки переносячи відповідні кінці на себе. Це ще ефективніше дозволяє вирішувати проблему супервитків при розплітанні ДНК.
г) Отже, «підтримуваний» топоізомеразами, фермент геліказа здійснює локальне розплітання подвійної спіралі ДНК на дві окремі нитки.
З кожною з цих ниток відразу зв'язуються спеціальні SSB -белки (від англ. Single Strand Binding Proteins; S на мал. 1.11). Останні мають підвищену спорідненість до одноланцюгових ділянок ДНК і стабілізують їх в такому стані.
Відмітимо: тим самим ці білки відрізняються від гістонів, які зв'язуються в першу чергу з дволанцюговими ділянками ДНК.
Ферменти полімеризації
а) Спеціальний білок виконує функції активатора праймази (АП на мал. 1.11). Після чого праймаза (П), використовуючи як матрицю відповідну ділянку одноланцюгової ДНК, синтезує коротку РНК-приманку, або праймер.
б) Далі в справу вступають ДНК-полімерази. У еукаріот відомо 5 різних ДНК-полімераз. З них β - і ε-полімерази беруть участь в репарації ДНК, γ-полімераза в реплікації мітохондріальної ДНК, а α- і δ-полімерази - в реплікації ядерної ДНК.
При цьому, по деяких припущеннях, α-полімераза пов'язана і з праймазой, і з δ-полимеразою, а остання, у свою чергу, з білком PCNA (від англ. Proliferating Cell Nuclear Antigen; P на мал. 1.11).
Цей білок виконує роль "прищепки", яка кріпить комплекс полімераз до реплікуємого ланцюга ДНК. Вважається, що в "застібнутому" стані він, як кільце, обхоплює ланцюг ДНК (мал. 1.13).
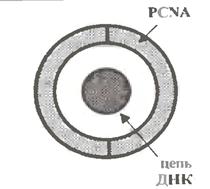
Тим самим попереджується передчасна дисоціація полімераз від даного ланцюга.
Зрозуміло, що ДНК-полімерази здійснюють послідовні включення дезоксирибонуклеотидів в ланцюг ДНК, що будується, - комплементарно нуклотидам батьківського ланцюга.
Але, крім того, ці ферменти, мабуть, мають і ряд інших важливих активностей. Для еукаріотичних ДНК-полімераз розподіл даних активностей ще не цілком ясно. Тому приведемо відомості відносно аналогічних бактеріальних ферментів.
У бактерій основну "роботу" по реплікації ДНК виконує ДНК-полімераза III, що має структуру димера. Саме з нею пов’язаний "зажим" типу білка PCNA.
Окрім ДНК-полімеразної активності, ДНК-полімераза III має ще одну - 3´→5'- екзонуклеазної. Остання спрацьовує в тих випадках, коли припустилась помилка і в ланцюг, що будується, включений "неправильный" нуклеотид. Тоді, розпізнавши дефект спаровування основ, фермент відщеплює із зростаючого (3´-) кінця останній нуклеотид, після чого знову починає працювати як ДНК-полімераза.
Таким чином, відбувається постійний контроль системи за результатом своєї діяльності.
в) Як ми знаємо, нові ланцюги ДНК утворюються спочатку у вигляді фрагментів - відносно коротких (фрагментів Оказаки) і дуже довгих. І кожен з них починається з праймерної РНК.
Коли ферментний комплекс, що рухається по батьківському ланцюгу, доходить до РНК-приманки попереднього фрагмента, "зажим", що зв'язує ДНК-полімеразу III з батьківським ланцюгом ДНК, розкривається, і цей фермент зупиняє роботу.
У дію вступає ДНК-полімераза I (як і раніше йдеться про бактеріальні ферменти). Вона приєднується до 3´-кінця зростаючого фрагмента (мал. 1.14).
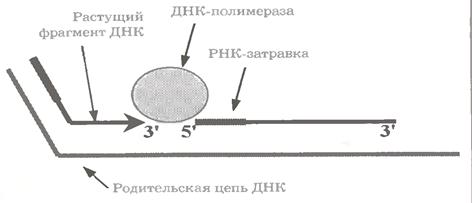
При цьому фермент вже не має стійкого зв'язку з цим фрагментом і з батьківським ланцюгом, та зате володіє навіть не двома, а трьома активностями.
Перша з них - "передня", або 5´ → 3´ - екзонуклеазна активність: послідовне відщеплення нуклеотидів з 5´-кінця РНК-приманки попереднього фрагмента.
На місце, що звільняється, фермент включає дезоксирибонуклеотиди, приєднуючи їх, як завжди, до 3'- кінця "свого" фрагмента (ДНК-полімеразна активність).
І, нарешті, подібно до ДНК-полімеразі III, він "не забуває" перевіряти і при необхідності коригувати свою діяльність - за допомогою "задньої", або 3´ → 5 '- екзонуклеазної активності, спрямованої на подовжуваний фрагмент.
Функція ДНК-полімерази I вичерпується, коли зростаючий фрагмент впритул доходить до дезоксирибонуклеотидів попереднього фрагмента.
Що стосується еукаріот, то тут функціональним аналогом бактеріальної ДНК-полімерази III є, мабуть, комплекс α- і δ-ДНК-полімераз; при цьому коригуюча 3´ → 5´ -екзонуклеазна активність властива δ-ДНК-полімеразі.
Функції ДНК-полімерази I теж розподілені між двома ферментами: 5' → 3'- екзонуклеазна активність (видалення РНК-приманки) здійснюється, ймовірно, спеціальною нуклеазою (Н на мал. 1.11), а ДНК-полімеразна активність (забудовування "проломів") - ДНК-полімеразою β (тій, що бере участь і в репарації).
г) Говорячи про ферменти полімеризації, не можна не сказати про найважчу з пов'язаних з ними проблем. Йдеться про синтез ланцюга ДНК, що запізнюється : як ми знаємо, напрям цього синтезу протилежний загальному напряму поширення реплікативної вилки.
Є, принаймні, дві гіпотези, що пояснюють ці протиріччя.
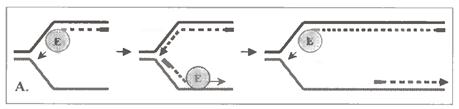
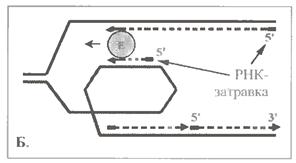
По одній з них (мал. 1.15, А), ферментний комплекс періодично припиняє утворення лідируючого ланцюга, переходить на другий батьківський ланцюг і синтезує черговий фрагмент Оказаки ланцюга, що запізнюється. Потім знову повертається на перший батьківський ланцюг і продовжує подовжувати лідируючий ланцюг ДНК, що будується.
За іншою версією (мал. 1.15, Б), на другому ланцюзі батьківської ДНК (матриці ланцюга, що запізнюється) в процесі реплікації формується петля. Тому напрям утворення фрагмента Оказаки на внутрішній ділянці петлі починає співпадати з напрямом руху полімеразного комплексу. Тоді останній може практично одночасно утворювати відразу обидва ланцюги ДНК - і що лідирує, і що запізнюється.
Можливо, з цим пов'язаний той факт, що бактеріальна ДНК полимераза III є димером, а у еукаріот α- і δ-ДНК-полімерази утворюють єдиний комплекс. Але і при такому механізмі ланцюг, що запізнюється, як неважко переконатися, не може утворюватись безперервно, а тільки у вигляді фрагментів.
Ферменти, що завершують реплікацію ДНК
В результаті дії усіх попередніх ферментів кожний новосинтезований ланцюг виявляється таким, що складається з фрагментів, що впритул примикають один до одного.
"Зшивання" сусідніх фрагментів здійснюється ДНК-лігазою (Л на мал. 1.11). Як і ДНК-полімерази, цей фермент утворює міжнуклеотидний (фосфодиефирний) зв'язок.
Але якщо в полімеразній реакції одним з учасників є вільний дНТФ (дезоксирибонуклеозидтрифосфат), то в ДНК-лигазній реакції обидва учасники - кінцеві дНМФ (дезоксирибонуклеозидмонофосфати) у складі "зшитих" фрагментів.
З цієї причини енергетика реакції інша, і вимагає зпряжений гідроліз молекули АТФ.
Відмітимо також, що ДНК-лігаза "зшиває" тільки такі одноланцюгові фрагменти, які знаходяться у складі дволанцюгової ДНК.
Але і це ще не усе. Молекула ДНК виявиться реплікованою не повністю, якщо не станеться спеціальний процес реплікації її кінців, або теломерних ділянок.
У цьому процесі ключову роль грає фермент теломераза, до якого останніми роками прикована увага багатьох дослідників. Тому розглянемо цей фермент і пов'язані з ним питання детальніше.
РЕПЛІКАЦІЯ ТЕЛОМЕРНИХ ВІДДІЛІВ ДНК
Основні уявлення
Суть проблеми кінцевої недореплікації
Уперше цю проблему сформулював А. М. Оловніков в 1971 р. Причому проблема відноситься тільки до лінійної ДНК.
Справа в тому, що ДНК полімеразна система, описана вище, залишає недореплікованими 3'- кінці материнських ланцюгів ДНК, тобто нові ланцюги виявляються укороченими (при дії тільки цієї системи) з 5 '- кінців.
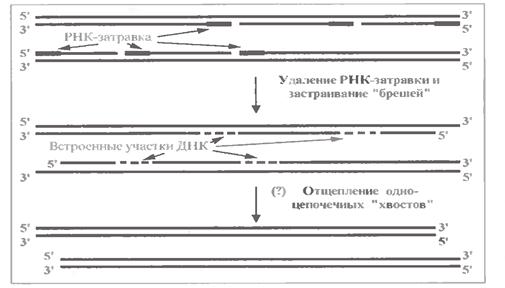
Це показано на мал. 1.16.
У кожному новому ланцюзі фрагмент Оказаки, що знаходиться біля 5'- кінця, як і зазвичай, починається з короткої РНК-приманки, яка (теж як завжди) потім видаляється спеціальною нуклеазою.
Але забудувати дезоксинуклеотидами "пролом", що утворюється, не може, оскільки будь-яка з ДНК-полімераз, як вже відзначалося, не здатна діяти "з нуля", а лише подовжує 3'-кінець вже наявного полінуклеотиду. Тут же такого кінца просто немає, чому і виходить, що новий ланцюг має бути дещо коротшим за старий.
Подібні кінці ДНК (де один ланцюг довший за інший) називають гострими, або оверхенгами.
Але, у свою чергу, гострі кінці навряд чи стабільні: вони, мабуть, атакуються екзонуклеазами, які понуклеотидно "відстригають" виступаючі одноланцюгові ділянки. Кінці ДНК стають "тупими". Хоча, за іншими даними, у людській молекулі ДНК зберігаються гострі кінці.
Як би то не було, за відсутності теломерази поділ клітини призводить до укорочення хромосом. З двох дочірніх хромосом одна коротша, ніж початкова, з однією теломерою, а друга - з протилежною.
Можна було б думати, що укорочення молекули ДНК, відповідно до довжини РНК-приманки, складає 10-15 нуклеотидних пар. Проте в реальності воно помітно більше (близько 50-65 н. п.). Ймовірно, це пов'язано з просторовими розмірами ДНК полімеразного комплексу: для його ефективного зв'язування при синтезі чергового фрагмента Оказаки необхідна ділянка батьківського ланцюга в декілька десятків нуклеотидів.
Середня кількість нуклеотидних пар в одній молекулі ядерної ДНК у людини - 120 млн. Таким чином, укорочення ДНК за відсутності теломерази складає за один клітинний поділ всього біля 0,00005 %. Здавалося б, це дуже мало.
Але якби в природі був зовсім відсутній механізм відновлення довжини теломер, то через осяжну кількість клітнних поділів хромосоми взагалі б зникли.
Таким чином, проблема кінцевої недореплікації хромосом має велике біологічне значення. Але виявляється, що вона прямо пов'язана і з такими "вічними" темами, як старіння і канцерогенез (про що буде сказано пізніше).
Буферні теломерні послідовності
В клітині вирішується проблема кінцевої недореплікації двома способами.
Один полягає в розташуванні на кінцях хромосомної ДНК спеціальних гексануклеотидних послідовностей, які не несуть генетичної інформації:
(5´) ЦТААЦЦ .. ЦТААЦЦ- - ГГТТАГ- -.ГГТТАГ (3´)
(3´) ГАТТГГ- - ГАТТГГ -ЦЦААТЦ-.. -ЦЦААТЦ (5´)
Неважко побачити, що ця схема виводить із структури лише один гексануклеотид:
(5´) ГГТТАГ (3´).
Його численні повтори розташовуються у 3´-кінца кожного ланцюга ДНК.
По складу цих повторів в кожній теломерній області ДНК розрізняють G- (чи Г -) ланцюг і С- (Ц- ) ланцюг. Очевидно, один і той же ланцюг ДНК в одній теломерній області виступає як G- ланцюг, а в іншій теломерній області - як С-ланцюг.
Всього теломерні відділи ДНК включають тисячі вказаних теломерних повторів. Їх загальна протяжність на одному кінці ДНК складає в клітинах ембріона людини 10-15 тисяч нуклеотидних пар. Таким чином, на обидві теломерні області доводиться близько 0,02 % від середньої довжини молекули ядерної ДНК людини (120 млн н. п.). У мишей теломерні послідовності довші - близько 100 тисяч н. п.
Як вже було сказано, теломерні повтори не несуть генетичної інформації. Тому якщо у відсутність теломерази відбувається втрата деякої частини цих повторів, це відображаеться на функціонуванні генома. У цьому-то і полягає, мабуть, основна роль теломер: своїм існуванням вони запобігають від недореплікації більш значущі області ДНК, тобто виконують роль своєрідного буфера.
Читайте також:
- Алгоритм формування комплексу маркетингових комунікацій
- АСУТП без керувального обчислювального комплексу.
- Гіпоопіка. Батьківське нехтування та неприйняття дитини може призвести до формування у неї комплексу неповноцінності через відчуття дитини себе небажаною.
- Гіпотези, які не відхиляються в експерименті, перетворюються на компоненти теоретичного знання про реальність: факти, закономірності, закони.
- Господарського комплексу
- ДЕРЖАВНЕ УПРАВЛІННЯ СИСТЕМОЮ ІНЖЕНЕРНО-ТЕХНІЧНОГО ЗАБЕЗПЕЧЕННЯ АГРОПРОМИСЛОВОГО КОМПЛЕКСУ
- Економічна оцінка ефективності роботи і розвитку пасажирського транспортного комплексу.
- Елементи й компоненти ландшафту
- Етапи і компоненти системного аналізу.
- Завдання комплексу маркетингових комунікацій
- Засоби маркетингового комплексу просування.
- Засоби маркетингового комплексу просування.
| <== попередня сторінка | | | наступна сторінка ==> |
| | | Подовження теломер за допомогою теломерази |
|
Не знайшли потрібну інформацію? Скористайтесь пошуком google: |
© studopedia.com.ua При використанні або копіюванні матеріалів пряме посилання на сайт обов'язкове. |