РЕЗОЛЮЦІЯ: Громадського обговорення навчальної програми статевого виховання
ЧОМУ ФОНД ОЛЕНИ ПІНЧУК І МОЗ УКРАЇНИ ПРОПАГУЮТЬ "СЕКСУАЛЬНІ УРОКИ"
ЕКЗИСТЕНЦІЙНО-ПСИХОЛОГІЧНІ ОСНОВИ ПОРУШЕННЯ СТАТЕВОЇ ІДЕНТИЧНОСТІ ПІДЛІТКІВ
Батьківський, громадянський рух в Україні закликає МОН зупинити тотальну сексуалізацію дітей і підлітків
Відкрите звернення Міністру освіти й науки України - Гриневич Лілії Михайлівні
Представництво українського жіноцтва в ООН: низький рівень культури спілкування в соціальних мережах
Гендерна антидискримінаційна експертиза може зробити нас моральними рабами
ЛІВИЙ МАРКСИЗМ У НОВИХ ПІДРУЧНИКАХ ДЛЯ ШКОЛЯРІВ
ВІДКРИТА ЗАЯВА на підтримку позиції Ганни Турчинової та права кожної людини на свободу думки, світогляду та вираження поглядів
- Гідрологія і Гідрометрія
- Господарське право
- Економіка будівництва
- Економіка природокористування
- Економічна теорія
- Земельне право
- Історія України
- Кримінально виконавче право
- Медична радіологія
- Методи аналізу
- Міжнародне приватне право
- Міжнародний маркетинг
- Основи екології
- Предмет Політологія
- Соціальне страхування
- Технічні засоби організації дорожнього руху
- Товарознавство продовольчих товарів
Тлумачний словник
Авто
Автоматизація
Архітектура
Астрономія
Аудит
Біологія
Будівництво
Бухгалтерія
Винахідництво
Виробництво
Військова справа
Генетика
Географія
Геологія
Господарство
Держава
Дім
Екологія
Економетрика
Економіка
Електроніка
Журналістика та ЗМІ
Зв'язок
Іноземні мови
Інформатика
Історія
Комп'ютери
Креслення
Кулінарія
Культура
Лексикологія
Література
Логіка
Маркетинг
Математика
Машинобудування
Медицина
Менеджмент
Метали і Зварювання
Механіка
Мистецтво
Музика
Населення
Освіта
Охорона безпеки життя
Охорона Праці
Педагогіка
Політика
Право
Програмування
Промисловість
Психологія
Радіо
Регилия
Соціологія
Спорт
Стандартизація
Технології
Торгівля
Туризм
Фізика
Фізіологія
Філософія
Фінанси
Хімія
Юриспунденкция
Фотосинтез.
Пластиди, їх функції і будова.
План.
Лекція: Пластиди, їх функції і будова.
Пластиди — це органели, властиві лише рослинним клітинам. Вони є гетерогенним сімейством органел, до якого входять пропластиди, хлоропласти, хромопласти, амілопласти, протеїнопласти та етіопласти.
У вищих рослин і деяких багатоклітинних водоростей пластиди утворюються з пропластид — дрібних тілець, які виявляють у меристематичних зонах. Це еліпсоїдні або сферичні структури діаметром 1–1,5 мкм. Залежно від місця розташування в рослині можуть формуватися пластиди різних типів.
Пропластиди — виступають в ембріональних клітинах промеристеми і меристеми. За будовою пропластиди нагадують мітохондрії, але відрізняються від них більшими розмірами і паралельним розміщенням внутрішніх мембран. Містять строму і дископодібні грани. Утворюються з недиференційованих зачатків пластид, які можуть інтенсивно ділитися. Спочатку вони круглі, згодом стають овальними. Це безбарвні, молоді стадії в розвитку всіх типів пластид.
Етіопласти — пластиди, характерні для листя, що виросло в темноті, а також для первинного листя та сім’ядолей проростаючого насіння. Їх можна розглядати як певні стадії розвитку хлоропластів. У етіопластах є багато компонентів хлоропластів, але не всі. Також у них немає справжніх тилакоїдів, а внутрішня мембрана утворює напівкристалічні структури — попередники тилакоїдів і ламел. Під дією світла етіопласти здатні швидко трансформуватися в хлоропласти.
Амілопласти — безбарвні пластиди, що не містять пігментів. Вони пристосовані для зберігання запасів крохмалю, і тому їх дуже багато в запасаючих органах — корінні, насінні, видозмінених стеблах, а також у клітинах кореневого чохлика. Крохмаль міститься безпосередньо в стромі амілопласту у вигляді зерен. В амілопластах бульб картоплі міститься по одному великому зерну діаметром до 100 мкм, а в амілопластах клітин кореневого чохлика таких зерен зазвичай вісім. Зрозуміло, що в запасаючих тканинах функція амілопластів пов’язана із синтезом, зберіганням і мобілізацією крохмалю (яка здійснюється в періоди потреби рослини у вуглеводах, наприклад під час проростання). А от у кореневому чохлику вони є структурами, що сприймають гравітацію.
Хромопласти — нефотосинтезуючі забарвлені пластиди, що містять переважно червоні, помаранчеві та жовті пігменти (каротиноїди). Найбільша кількість хромопластів міститься в плодах (наприклад, томата й червоного перцю), у квітках, яскраве забарвлення яких приваблює комах і птахів, що сприяє запиленню рослин і поширенню насіння, а також у коренеплодах (наприклад, моркви). Найчастіше хромопласти розвиваються з хлоропластів і мають такі самі форму та розміри. Ці органели мають різну внутрішню будову залежно від виду рослини або тканин, у яких вони наявні.
Хлоропласти — зелені фотосинтезуючі пластиди . У вищих рослин вони локалізуються переважно в стовпчастій і губчастій паренхімі листя. У клітинах зелених тканин усіх інших видів також містяться хлоропласти. Кількість хлоропластів у клітині варіює в досить широких межах: від одного великого в багатьох одноклітинних водоростей (Chlamidomonas, Chlorella) і печінкових мохів (Anthoceros) до 300–400 у клітинах палісадної паренхіми листя покритонасінних рослин. Різними є також розміри та форма хлоропластів. У вищих рослин найчастіше вони є двоопуклими еліпсоїдами розміром 3–10 мкм. Згадуваний Anthoceros має єдиний хлоропласт діаметром 40 мкм. Найбільш різноманітні форми хлоропластів виявлено у водоростей: у Spirogyra — спіральні, у Euglena — сочевицеподібні, у Zygnema — такі, що нагадують за формою зірки, а в Oedogonium хлоропласти формують сітчасту структуру.
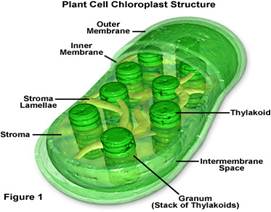
Будова хлоропласта
Усі хлоропласти містять зелений пігмент хлорофіл, проте колір їх не завжди зелений. У деяких диких форм (бурі та червоні водорості) і культурних декоративних рослин (колеус, драцена, бегонія) їх зелене забарвлення відтіняється іншими пігментами. Головною функцією хлоропластів є фотосинтез. Світлові реакції фотосинтезу пов’язані із системою внутрішніх мембран хлоропластів. Саме в них міститься хлорофіл. Уся система мембран складається з безлічі плоских мішечків — тилакоїдів.
Тилакоїди утворюють скупчення — ґрани, схожі на купки монет. Ґрани сполучаються між собою плоскими одиночними шарами — ламелами. Внутрішній вміст хлоропласта називають стромою. У стромі відбуваються темнові реакції фотосинтезу. У ній міститься багато ферментів, ліпіди, цукри. Крохмаль накопичується в стромі у вигляді яйцеподібних зерен діаметром до 1,5 мкм, розташованих поблизу тилакоїдних мембран.
Походження, онтогенез і взаємоперетворення пластид на сьогодні можна уявити таким чином. В еволюційному сенсі первинним вихідним типом пластид є хлоропласти. Із них у ході розчленовування тіла рослин на органи виникли пластиди решти типів. У процесі індивідуального розвитку (онтогенезу) пластиди майже всіх типів можуть перетворюватися одна на одну. Найбільш звичайними процесами є перетворення етіопластів на хлоропласти (розвиток листя в бруньці, розвиток зародка із заплідненої яйцеклітини) і хлоропластів на хромопласти (пожовтіння листя восени, змінення забарвлення плодів під час дозрівання). Перший процес відбувається завдяки поступовому утворенню в етіопластах характерної для зелених пластид внутрішньої мембранної системи (що включає ґрани), на якій відбувається синтез хлорофілу. Деякі вчені називають етіопласти ембріональних (таких, що діляться) клітин ростової бруньки протопластидами (тобто зачатковими пластидами). Проте вивчення цих утворень показує, що вони є звичайними дрібними етіопластами.
Процес перетворення хлоропластів на хромопласти полягає в зменшенні розмірів пластид, поступовому руйнуванні внутрішніх мембран (тилакоїдів і ламел) і накопиченні речовин у пластоглобулах, число та розмір яких збільшуються. Урешті-решт хлорофіл повністю руйнується та перестає маскувати каротиноїди, які тепер виразно виступають і зумовлюють жовте, жовто-гаряче й червоне забарвлення осіннього листя. Переважним компонентом пластид стають пластоглобули. У природних умовах зворотний процес не відбувається: хромопласти можна розглядати як кінцевий етап розвитку пластид (етап старіння).
У хромопласти можуть перетворюватися й амілопласти (наприклад, у деяких видільних тканинах у процесі їх старіння). якщо під час перетворення хлоропластів в етіопласти, яке може відбуватися, поранити рослину або помістити її в темноту, внутрішня мембранна система також значною мірою руйнується, хлорофіл зникає, але накопичення пластоглобул не відбувається. Цей процес є оборотним. Так, якщо помістити рослину на світло, з етіопластів знову розвиваються хлоропласти.
У процесі розвитку клітини пластиди виникають лише з пластид, а не з інших структур. Чисельність їх у клітині збільшується за рахунок поділу шляхом утворення перетяжок, услід за якими відбувається ріст дочірніх органел.
Фотосинтез і хемосинтез.
| Рік відкриття | Учений | Вклад у розвиток науки |
| 1674 р. | А. ван Левенгук | Уперше спостерігав хлорофіл |
| 1753 p. | М. Ломоносов | Рослини поглинають з повітря «жирну вологу», яка є їх їжею і перетворюється на «тіло рослин» |
| 1771 р. | Дж. Прістлі | Вивчав повітряне живлення рослин. Визначив, що гілочка м’яти під ковпаком здатна зберегти життя миші |
| 1779 р. | Я. Інгенгауз | Довів, що «поліпшення повітря» рослинами відбувається лише під впливом світла, а в темряві вони подібно до тварин «погіршують повітря» |
| 1783 р. | Ж. Сенеб’є | Довів, що виділення на світлі « чистого повітря» відбувається тільки за наявності «зіпсованого» |
| 1840 р. | Ж. Б. Бусенге | Кількісно довів, що рослини поглинають з повітря вуглекислий газ і виділяють кисень |
| 1840 р. | Р. Майєр | Кількісно довів, що рослини поглинають з повітря вуглекислий газ і виділяють кисень |
| 1840 р. | Р. Майєр | Рослиний світ утворює резервуар, в якому накопичуються сонячні промені |
| 1854 р. | Г. Гельмгольц | Уявлення про роль рослин у накопиченні енергії сонячного світла ще не доведені, це поки лише гіпотеза |
| 1871—1875 pp. | К. А. Тимірязєв | Довів, що рослина не випадково зелена, з цим забарвленням пов’язана її здатність використовувати сонячну енергію для засвоєння вуглекислого газу і синтезу органічних сполук |
| 1877 р. | К. А. Тимірязєв | Процес перетворення сонячної енергії з допомогою хлорофілу і з участю вуглекислоти і води на хімічну потенціальну енергію був названий фотосинтезом |
| 1940 р. | Х. Фішер | Встановив структуру хлорофілу |
| 1960 р. | Р. В. Вудворт | Здійснив синтез хлорофілу |
Фототрофи використовують для синтезу органічних сполук енергію світла. Процес утворення органічних сполук із неорганічних завдяки перетворенню світлової енергії в енергію хімічних зв'язків називають фотосинтезом.
До фототрофних організмів належать зелені рослини (вищі рослини, водорості), деякі тварини (рослинні джгутикові), а також деякі прокаріоти — ціанобактерії, пурпурові та зелені сіркобактерії.
Основними з фотосинтезуючих пігментів є хлорофіли. За своєю структурою вони нагадують гем гемоглобіну, але в цих сполуках замість заліза присутній магній. Залізо потрібне рослинним організмам для забезпечення синтезу молекул хлорофілу (якщо в рослину залізо не надходить, то в неї утворюються безбарвні листки, нездатні до фотосинтезу).
Більшість фотосинтезуючих організмів має різні хлорофіли хлорофіл а (обов'язковий), хлорофіл Ь (у зелених рослин), хлорофіл с (у діатомових і бурих водоростей), хлорофіл d (у червоних водоростей). Зелені й пурпурові бактерії містять особливі бактеріохлорофіли.
В основі фотосинтезу лежить окиснювально-відновний процес, пов'язаний із перенесенням електронів від сполук постачальників електронів (донорів) до сполук, які їх сприймають (акцепторів), з утворенням вуглеводів і виділенням в атмосферу молекулярного кисню.
Світлова енергія перетворюється на енергію синтезованих органічних сполук (вуглеводів) в особливих структурах - реакційних центрах, що містять хлорофіл а.
У процесі фотосинтезу у зелених рослин і ціанобактерій беруть участь дві фотосистеми — перша (І) та друга (II), які мають різні реакційні центри та пов'язані між собою через систему перенесення електронів.
Кожна із фотосистем містить дві–три сотні молекул пігментів, що уловлюють світло різної довжини хвилі. Ці пігменти утворюють світлозбиральний комплекс, що збільшує площу поверхні поглинання світла та ефективність його використання для фотосинтезу. Зрештою поглинене пігментами світло передається на особливу молекулу хлорофілу в кожній із фотосистем.
Кінцевою молекулою хлорофілу у ФС I є Р700 (від лат. pigmentum — фарба, а цифри означають довжину хвилі максимуму поглинання в нанометрах), у ФС II — Р680. Як бачимо, обидва піки поглинання розміщені в червоній області спектра, саме тому практично всі наземні рослини та мілководні водорості є зеленими.
ФС I містить близько 200 молекул хлорофілів, 50 молекул каротинів і декілька цитохромів, а ФС II — також близько 200 молекул хлорофілів, 50 молекул ксантофілів, цитохроми та шість йонів Манґану. Крім пігментів кожна фотосистема містить комплекс окиснювально-відновних компонентів, що є донорами й акцепторами електронів. Усі компоненти фотосистем організовані в електронтранспортний ланцюг — систему асоційованих з тилакоїдною мембраною переносників електрона.

Світлозбиральнийкомплекс
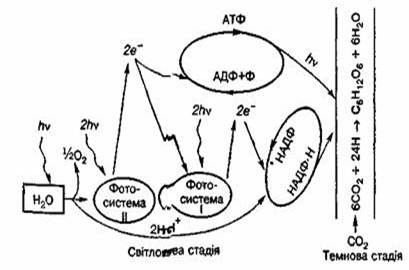
Процес фотосинтезу відбувається в дві фази — світлову та темнову.
Світлову фаза - реакції перебігають у мембранах особливих структур хлоропластів - тилакоїдів за наявності світла- фотосинтезуючі пігменти вловлюють кванти світла (фотони). Поглинання фотонів приводить до «збудження» одного з електронів молекули хлорофілу, який за допомогою молекул — переносників електронів переміщується на зовнішню поверхню мембрани тилакоїдів, набуваючи певної потенційної енергії.
У фотосистемі І цей електрон може повертатись на свій енергетичний рівень і відновлювати її, а може передаватись такій сполуці, як НАДФ (нікотинамідаденіндинуклеотидфосфат). Електрони, взаємодіючи з іонами водню, які є в навколишньому середовищі, відновлюють цю сполуку:
НАДФ⁺ + 2е+2Н⁺→НАДФ Н₂
Нагадаймо, що коли певна сполука віддає електрон — вона окиснюється, а коли приєднує — відновлюється. Відновлений НАДФ (НАДФ • Н2) згодом постачає водень, потрібний для відновлення атмосферного СО2 до глюкози (тобто сполуки, в якій запасається енергія).
Подібні процеси відбуваються й у фотосистемі II. Збуджені електрони, повертаючись на свій енергетичний рівень, можуть передаватись фотосистемі І і таким чином п відновлювати. Фотосистема II відновлюється за рахунок електронів, які постачають молекули води. Під дією світла за участю ферментів молекули води розщеплюються (фотоліз води) на протони водню та молекулярний кисень, який виділяється в атмосферу, а електрони використовуються на відновлення фотосистеми II:
2Н₂О-→4Н⁺+О₂+2е
Енергія, вивільнена при поверненні електронів із зовнішньої поверхні мембрани тилакоїдів на попередній енергетичний рівень, запасається у вигляді хімічних зв'язків молекул АТФ, які синтезуються під час реакцій в обох фотосистемах. Деяка її частина витрачається на випаровування води. Таким чином, під час світлової фази фотосинтезу утворюються багаті на енергію (яка запасається у вигляді хімічних зв'язків) сполуки: синтезується АТФ і відновлюється НАДФ. Як продукт фотолізу води в атмосферу виділяється молекулярний кисень.
4hν + Н2О + НАДФ+ + 2АДФ + 2Фн → НАДФН + Н+ + 2АТФ + ½О2,
де hν — фотони світла;
Фн — молекула ортофосфатної кислоти.
Як видно з рівняння, для фотолізу однієї молекули води, синтезу двох молекул АТФ, відновлення одного НАДФ+ і виділення півмолекули кисню потрібна енергія чотирьох квантів світла. Варто зазначити, що весь кисень, що виділяється під час фотосинтезу, походить із води, а не з вуглекислого газу
| <== попередня сторінка | | | наступна сторінка ==> |
| Співпраця та безпека в НАТО. | | | Темнова фаза |
|
Не знайшли потрібну інформацію? Скористайтесь пошуком google: |
© studopedia.com.ua При використанні або копіюванні матеріалів пряме посилання на сайт обов'язкове. |