РЕЗОЛЮЦІЯ: Громадського обговорення навчальної програми статевого виховання
ЧОМУ ФОНД ОЛЕНИ ПІНЧУК І МОЗ УКРАЇНИ ПРОПАГУЮТЬ "СЕКСУАЛЬНІ УРОКИ"
ЕКЗИСТЕНЦІЙНО-ПСИХОЛОГІЧНІ ОСНОВИ ПОРУШЕННЯ СТАТЕВОЇ ІДЕНТИЧНОСТІ ПІДЛІТКІВ
Батьківський, громадянський рух в Україні закликає МОН зупинити тотальну сексуалізацію дітей і підлітків
Відкрите звернення Міністру освіти й науки України - Гриневич Лілії Михайлівні
Представництво українського жіноцтва в ООН: низький рівень культури спілкування в соціальних мережах
Гендерна антидискримінаційна експертиза може зробити нас моральними рабами
ЛІВИЙ МАРКСИЗМ У НОВИХ ПІДРУЧНИКАХ ДЛЯ ШКОЛЯРІВ
ВІДКРИТА ЗАЯВА на підтримку позиції Ганни Турчинової та права кожної людини на свободу думки, світогляду та вираження поглядів
- Гідрологія і Гідрометрія
- Господарське право
- Економіка будівництва
- Економіка природокористування
- Економічна теорія
- Земельне право
- Історія України
- Кримінально виконавче право
- Медична радіологія
- Методи аналізу
- Міжнародне приватне право
- Міжнародний маркетинг
- Основи екології
- Предмет Політологія
- Соціальне страхування
- Технічні засоби організації дорожнього руху
- Товарознавство продовольчих товарів
Тлумачний словник
Авто
Автоматизація
Архітектура
Астрономія
Аудит
Біологія
Будівництво
Бухгалтерія
Винахідництво
Виробництво
Військова справа
Генетика
Географія
Геологія
Господарство
Держава
Дім
Екологія
Економетрика
Економіка
Електроніка
Журналістика та ЗМІ
Зв'язок
Іноземні мови
Інформатика
Історія
Комп'ютери
Креслення
Кулінарія
Культура
Лексикологія
Література
Логіка
Маркетинг
Математика
Машинобудування
Медицина
Менеджмент
Метали і Зварювання
Механіка
Мистецтво
Музика
Населення
Освіта
Охорона безпеки життя
Охорона Праці
Педагогіка
Політика
Право
Програмування
Промисловість
Психологія
Радіо
Регилия
Соціологія
Спорт
Стандартизація
Технології
Торгівля
Туризм
Фізика
Фізіологія
Філософія
Фінанси
Хімія
Юриспунденкция
Ісіктер иммунотерапиясы.
Қатерлі ісікті иммунотерапиясының ішіндегі айқын бір әдіс иммун жүйесінің арнайы стимуляциясына негізделген.
1. Лимфокин арқылы активтелген аутологиялық Т-лимфоциттер/киллерлер колданылады. Ол үшін науқастың лимфоциттерін организмнен тіс ИЛ-2 ж.т.б. цитокендермен активтейді.
2. Басқа бығытта – ісікке қарсы вакцинаны жасау. Осы үшін дендритті клеткаларды ісікті арнайы антигендерімен байланыстырады немесе оларға осы антигендерді кодтайтын ДНК-ны еңгізеді.
3. СД8 Т-клеткалардың цитотоксикалық әсерін күшейту әдісі ісікті клеткалардың геномына, цитокиндерді кодтайтын ДНК-ның бөлігін енгізуіне негізделеді
Сонымен қатар онкологиялық аурулардың иммунотерапиясында фармакологиялық препараттармен, токсиндермен жэне радиозаттармен коньюгацияланған моноклоналды антигендер қолданылады.
Ісікке қарсы иммунитетті құрудың басқа әдісі иммунды жауапты антигендер арқылы арнайы емес стимуляциясына негізделген. Әдетте, стимуляторлар макрофагтарға әсер етеді. Осы топтар ішіндегі препараттарға БЦЖ вакцинасы жатады. Вакцинаны ісіктер ішіне енгізу, ісіктің толық регрессиясын шақырады немесе ісіктің өсуін тежейді. Клиникада меланоманы емдеу кезінде БЦЖ қолдану белгілі жетістіктерге әкелді. СД8 Т-клеткаларының цитотоксикалық эффекторларға дейін пісіп жетілуі пептид+МНС І класс молекула комплексін танудан басқа антиген танушы клеткалардың В7 космтимуляторы әсерінен тұрады. Иммунотерапияда ИНФ-гамма, ИЛ2, ФНО қолданылады.
HLA-жүйесі
Генетикалық бөгде тіндердін тұрақтамауна жаупты негізгі антигендерге – гистосәйкестік антигендері жатады, оларды кодтайтын гендерін - гистосәйкестік гендері деп атайды. Олар өздерінің антигендігін өзінің емес, генетикалық бөгде организмнің иммун жүйесімен анықталғанда ғана көрсетеді, мысалы мүшелерді аллотрансплантациялау кезінде.
Ұқсас антигендері бар тіндерді - гистосәйкестік тіндер деп атайды. Бул тіндер тұрақтамаушылық реакциясына келтіретін иммундық жауапты шақырмайды. Манызды антигендік айырмашылықтары бар - гистосәйкессіз болып келеді, олар тұрақтамаушылық реакциясын шақыратын иммундық жауапка әкеледі.
Гистосәйкестікті орнататын антигендер 40 -тан астам әртүрлі локустарда шифрленген., бірак көрнікті тұрақтамаушылық реакциясына жауапты локустар гистосәйкестіліктің бас –МНС –комплексінде орналасады.
Бул комплекс барлық сүткөректілерде және құстарда анықталған. Адамдарда HLA-жүйесі (Human Leukocyte Antigens) деп аталған. HLA-жүйесінің гендері трансплантациялық (тін сәйкесті) антигендердің синтезің бақылайды. Трансплантациялық антигендер ең көп мөлшерде лимфоциттерде,теріде; аздауөкпе, бауыр, ішек, жүректе; ең аз – ийда болады. Жалпы, олар ядролы клеткалар бетінде және тромбоциттерде табылады. Донор мен реципиенттің трансплантациялық антигендер бойынша толық сәйкестілігі моногзиготалы егіздерде ғана болады. Ал басқа жағдайда тұрақтамаушылық реакциясын шақырады.
HLA-жүйесіадамның 6 хромосомасының қысқа иығында орналасады, ғ локустан тұрады:А,В,С, Д, ДR. Әр локусқа гендердің көпшілік аллельдік түрлері кіреді. А-локусына 23,локусына- В- 49, С-локусына- 8 аллельдер кіреді. Ал Д- локус өзі 2 локустан турады: DG,DP. DG- локусына “ аллель кіреді, DG-і,DP-, аллельдер анықталған.
HLA-комплексіне үш гендер класстары кіреді.
1 класс гендеріне АВС локустар іреді. Олар антидене түзілуін, цитотоксикалық Т-лимфоциттерді, Т-супрессорларды стимульдейтін, трансплантациялық антигендердін синтезін бақылайды. HLA-1 класс антигедері организмнің барлық ядролы клеткаларда , Т-лимфоциттерде экспрессияланған және өзіндік тануда манызды роль атқарады.
2 класс гендеріне иммундық жуаптың гендері (IR) жатады. HLA- 2 класс анти- гендері В-лимфоциттерде, макрофагтарда, дендритті клеткаларда , активтелген Т-лимфоциттерде орналасады. HLA-2 класс антигендері иммунокомпетентті клеткалардың әсерлесүіне , антигендерді таныстыруында қатысады.
3 класс гендері С2,С4 – комплементтің компонентері, Вf –пропердин жүйесінің В-фаторы түзілуің және В-лимфоциттердегі С3 рецепторының экспрессиясын бақылайды
НLA-А және В локустарының антигендері күшті трансплантациялық антигендер қатарына жатады. Пратикада трансплантат күшті антиген бойынша біреуі ғана сай келмесе, донор мен реципиент оптималды болып саналады.
ТРАНСПЛАНТАЦИЯ ИММУНОЛОГИЯСЫ
Трансплантатты қабылдамау реакциясы
Тінді немесе мүшені генетикалық ерекшелігі басқа индивидумға салған кезде салынған биологиялық материалдың ажырау реакциясы байқалады. Тұрақтамаушылық реакциясы МНС-комплекс пен кіші гистосәйкестік комплексіне жататын қан тобының антигендеріне қарсы жүреді. МНС адамдарда - HLA-жүйесі деп аталған антигендері бойынша донор мен реципиент сәйкес болмаса, тұрақтамаушылық реакциясы жүреді. Трансплантаттың біріншілік ажырау уақыты – 14 күн. Екіншілік трансплантат одан жылдам ажырайды: 5-7 күнде. Трансплантаттық иммунитеттің даму реакциясы 3 этаптан тұрады:
1. трансплантаттың бөгде антигендерін тану;
2. транслпантатты ажырау реакциясының эффекторлары жетіліп жиналуы;
3. трансплантаттың ыдырауы.
Бірінші этапта трансплантат антигендерін цитотоксикалық Т-лимфоциттер (СД8), хелперлер және қабынулық Т-клеткалардың ізашарлары мен (ТнО) танудан басталады. Танығаннан кейін аймақтық лимфоидты тінге көшеді. Кейбір донордың лейкоциттері реципиенттің аймақтық лимфоидты түйіндеріне көшеді да бөгде зат болып танылады және реципиенттін лимфа түйіндеріндегі лимфоциттерді активтейді№ Сонымен олар трансплантат тұрақтамауна күшті әсер етеді.
Осы жерде 2 этап басталады. Тұрактамаушылық реакциясында ен манызды қызметті баяу жүретін аса сезім талдық реакциясың қатыстыратын клеткалық реакциялар және цитотоксикалық Т-клеткалар аткарады. СД8 ізашарлары жетілген цитотоксикалық Т-клеткаларына аусады. Бос трансплантатты антигендер лимфоидты тінге түсіп, антигентаныстырушы клеткалармен жұтылады және Тн1 және Тн2 клеткаларын жауапқа қосады. Танудың нәтижесі болып Тн1-ң хемотоксикалық факторы антиген енген аймаққа макрофагтарды және қабынудың басқа клеткаларын шақырады. Антигентаныстырушы, В-клеткалар, Тн2 клеткалар қосылып қабылдамау реакциясының қосымша тізбегі ретінде гуморалды иммунды жауап түзеді. Натуалды киллерлер Тн1--лимфоциттердің цитокиндрінің әсерінен активтенеді.
3 этапта бөгде тіннің ажырауы болады. Бул процесс жетілген СД8 Т-клетканың, Тн1 көмегімен белсендірілген макрофагтардің, иммуноглобулиндер және цитокиндермен белсендірілген нағыз киллерлердің көмегімен іске асды.
Бөтен тіндердің СД8 клеткалардың цитотоксикалы киллерлік әсері апрқылы лизисі бірнеше кезеңдерден тұрады.
1 кезең – СД8 Т-клеткалардың беткей бөгде антигендерімен байланысуы (трансплантатты вирустың пептиді, рак антигендері).
2 кезең – “летальды соққы”. Негізгі жағдай болып табылады. Клетка нысананың өлуін анықтайды. Эффектормен клетка нысана механикалық бірігуі оларды өлімнен сақтамайды. Бұл этапта клетка мембранасының өткізгіштігі артады, натрий-калий насосының балансы бұзылады. “Летальды соққы” механизмі әлі толық зерттелмеген. Клетка мембранасын зақымдайтын токсиндердің бірі – лимфотоксин (ісіктердің некрозының β-факторы).
3-кезең – нысана клетка лизисі болады. бұл зақымдалған мембрана арқылы Н2О
көп мөлшерде өтеді. Осы процестің нәтижесінде “нысана клетка” мембранасы жыртылады және ол өледі. Эффекторлы клетка сақталады және ары қарай цитолитикалық қасиетке ие болады.
Трансплантацияның зандары:
1. аутотрансплантатта, изотрансплантатта донормен реципиенттің генетикалық жақындығы себебінең істен шықпайды.
2. Аллогенді және ксеногенді трансплататтар әр-кашан тұрақтамаушылыққа ұшырайды.
3. Тұрақтамаушылықтың иммундық механизмдері иммунологиялық ерекшелігі мен естелігіне байланысты.
4. Жедел тұрақтамаушылық реакциясын бас гистосәйкестілік комплексі бақылайды.
5. Бірінші ұрпақты гибридтер- F1 ата-анасының уекеуінен де салынған трансплататтың тұрақтамауын шақырмайды, бірақ ата-анасынынң әр-біреуі гибридтен салынған трансплантаттың тұрақтамауын шақырады.
Жеделдеу тұрақтамаушылық реакциясы 24 сағаттың арасында өтуі мүмкін. Негізінде -аниденелер арқылы комплементтің активтенуі салынған тіннің васкуляризациясына кедергі етіп трансплантат жылдам істен шығады.
Жедел тұрақтамаушылық реакциясы әдетте трансплантациядан кейін бірінші апталар ішінде өтеді (7-14 шакты күннің ішінде). Ол клетка-тәуелді, қабыну Т-клеткалардың (Тх1) активтенуі арқылы цитокиндердің бөлінуі және макрофагтар мен лимфоциттердің зақымдану ошагында жиналуы.
Созылмалы тұрақтамаушылық реакциясы трансплантациядан кейін айлар, жылдар өткенде болуы мүмкін. Механизміне гуморалды және клеткалық реакциялар кіреді. Созылмалы тұрақтамаушылық реакциясы қайталанған трансплатацияны талап етеді.
Сонымен, мүшелер мен тіндерді аустырып салған кезде донор мен реципиентті гистосейкестік дәржесі бойынша іріктеу кажет. Біріншіден, донор мен реципиентте АВО-антигендері бойынша топтық сәйкестілігін анықтайды. Донор мен реципиенттін HLA-антигендерін серологиялық әдіспен іріктеу үшін микролимфоциттітоксикалы тест (МЛЦТ) қолданады.
Кәзірге кезде гистосейкестік антигендерді полимеразды тізбектік реакция (РСR) көмегімен анықтауға болады.
Трансплантат “иесіне қарсы” реакциясы
Туа біткен және жүре пайда болған иммундық жетіспеушілікті компенсациялау үшін клиникалық практикада қан түзу және лимфоидты тін клеткаларын алмастыруға тура келеді. Клеткалық трансплататта иммунокомпетентті клеткалар болады. Сол себепті реципиент клеткаларында реакциялар дамиды. Енгізілген бөгде лимфоциттері генетикалық ерекшеліктеріне байланысты реципиенттің туыс емес антигенің танып, трансплантат “иесіне қарсы антиген спецификалық реакцияны қалыптастырады. Тану процессіне донордың сүйек кеминдегі лимфоциттердің 2 субпопуляциясы қатысады: СД8 Т-клеткаларының изашарлары (цитотоксикалық Т-клетка) және СД4 Т-клетка. Бұл реакцияның нәтижесінде жетілген СД8, СД4-Тн1-клеткалармен түзілетін цитокиндер, хемотоксикалық факторлар арқылы активтенген макрофагтар жиналып реципиенттің тіндерін шабуылдайды. Көк бауырда немесе лимфа түйіндерде клеткалардың саны енгізілген лимфоциттің пролиферациясынан ғана емес, сонымен қатар реципиенттің клеткаларының қатысуына байланысты.
Сонымен, трансплантат “иесіне қарсы” реакциясын алдын алу үшін қажет:
1. донормен реципиентті HLA-антигендері бойынша, оның ішінде күшті трансплантациялық антигендермен бірге, DR-антигендері бойынша, іріктеу.
2. Трансплантаттан жетілген Т-лимфоциттерді алыстату
3. Иммунодепрессантты препараттарды колдану
Жоғары сезімталдық реакциясы.
Антигендердің белгілі бір формалары ағзамен қайталанып, қатынасқа түскенде өз негізінде арнайы реакция шақыруы мүмкін, бірақ жедел қабынулық жауаптың арнайы емес клеткалық және молекулалық факторларын өзіне қосады. Жоғарлаған рекативтіліктің екі формасы белгілі: жедел және баяу типті асасезімталдық. Реакцияның I-типі Jg E антиденесінің қатысуымен айқындалады, олар базофильдер мен мес клеткаларға цитрофильді болып келеді, сонымен қатар JgG қатысуымен. Екнші тип қабыну аймағында макрофагтардың жиналуын қамтамасыз ететін, реакцияның негізгі эффекторлары ретінде қабынудың Т- клеткалар көмегімен жүзеге асырылады (Тн1; бұл клеткалардың басқа белгілеуі- Тбст).
II-шi типті асасезімталдық клетка бетінде орналасқан малекулалардың JgG антидемен байланысуына негізделген. Жоғары сезімталдықтың бұл типі кейде дәрілік заттарды ұзақ уақыт қолданғанда дамиды. Мысалы, квинидин- жүрек аритмиясы кезінде қолданылатын препарат, эритоциттердің бетінде сорбирленуге қабілетті. Алдыңғы немесе индуцирленген JgG арнайы антиденелері болғанда, эритоциттер бетінде осы антиденелердің препаратпен әсерлесуі жүреді. Реакцияға комплемент қосылуы нәтижесінде эритроидты клтеткалардың көптеп өлуі байқалады. Осы асасезімталдықтың екінші типі сирек кездеседі.
Аса сезімталдықтың ІІІ- типі JgG ергіш антигенмен әсерлескенде жүзеге асырылады. Антиген антиденемен агрегаттары (иммунды кешендер) белгілі бір тінді аймақтаында орналасқан кезде патологиялық жағдай дамиды. Иммунды комплекстер кез- келген гуморальді жауап кезінде түзіледі. Бірақ олар кез келген жағдайда патологияның патогенетикалық фактор болмайды және бұл ең алдымен олардың өлшемдерімен байланысты. Жеткілікті ірі комплекстер комплементпен әсерлескеннен кейін фагоциттермен жұтылып, ағзадан шығарылады.
Осы уақытта ұсақ комплекстер, антигеннің көпшілігінде түзілетін, тамырлар қабырғаларында жиналуы мүмкін және тамырлық зақымданудың алғашқы себебі ретінде болуы мүмкін. Ағзада JgG антиденні болғанда, арнайы антиген теріге енгенде, Артюс реакциясы деп ататын локальді қабыну процесі дамиды. Тін қабаттарында түзілетін комплекс комплемент жүйсін белсендіреді, нәтижесінде қабыну медиаторлары- С5а, С3а- компонеттері жиналады. Олар өз кезегінде локальді қабыну жауабының басталуын қамтамасыз етеді, тамырлардың өткізгіштігін күшейтеді. Қабыну ошағында нейтрофильдер, тромбоциттер жиналады, сұйықтың көлемі ұлғаяды. Кез келген басқа қабыну жауабы сияқты, Артюс реакциясының да қорғаныс сипаты бар. Ол антигеннің ішкі мүшелерге енуіне кедергі жасайды.
Интактты организмге бөтен белокты көп мөлшерде енгізгенде, сарысулық ауру- жоғарғы сезімталдықтың ерекше формасы ретінде, дамуына әкеледі. Қазіргі кезде жылқының арнайы антисарысуын улы жыландардың шағуына шалдыққандарға қолданады. Бірақ, енгізілген сарысудың белоктарына иммунизация арқылы асқынулар пайда болуы мүмкін. Антисарысу инъекциясы нәтижесінде бөтен белоктарға қарсы JgG антиденелері түзіле бастайды. Түзілген антиденелер инъекциядан қалған белокты антигендермен байланысып, антиген- антидене комплекстерді түзеді. Комплекстің патогенетикалық әсері дене температурасының көтерілуімен, безгек, есекжем, тері қышуы, және протеинуриямен көрінеді.
JgЕ- мен шақырылған жоғарғы сезімталдық үшін, JgG-мен байланысты аурулардың әр түрлі формаларының дамуында антигеннің ағзаға ену әдісі мен дағдысының критикалық
маңызы бар.
Аллергиялық реакциялардың медиаторлары.
Шұғыл типті жоғарғы сезімталдық реакциясына жауапты химиялық медиаторлардың екі негізгі класы бар. Алдыңғы, немес біріншілік, медиаторлар молекулалар болып табылады, олар базофилдер мен бұлтты клеткалар түйіршектерінде жиналып қойған, және клетканың антигенмен қатынасынан кейін экстрацеллюлярлы ортаға секреттеле бастайды.
Бұл медиаторлар молекулалардың төрт негізгі түрлерімен көрсетілген:
1)вазоактивті аминдермен- гистамин, серотонин.
2) гранулоциттер үшін- хемотоксикалдық факторлармен.
3) энизимдермен.
4) протеогликандармен- гепарин (бұлтты клеткаларда) және хондроинтисульфат (базофилде).
Антиген антиденемен байланысқаннан кейін бұлтты клеткалар және базофилдермен de-novo синтезделетін молекулалар екіншілік медиаторлар болып табылады. Негізінен екіншілік медиаторлар лейкотриен мен тромбоциттерді белсендіруші факторларды қосатын липидтердің өнімдері ретінде ұсынылады.
Аллергиялық зақымданулардың басты медиаторының- гистаминнің нысанасы тегіс бұлшықеттер, қантамырлары, кейбір экзокринді бездер, лейкоциттер болып тбылады. Аллергиялық реакциялардың әр түрлі дамуы бірнеше сатыдан тұрады. Сенсибилизденген организмде арнайы JgЕ- антиденелері бұлтты клеткалардың бетінде бар. Сол аллерген ағзаға қайталап, енгенде JgЕ- мен әсерлеседі. Аллергеннің JgЕ- мен байланысуы клетка ішіне Са2+ енуін қамтамасыз етеді, нәтижесінде клетка белсендіріледі және клетка ішілік гранулдардан гистами босатылады. Медиатор нысана клеткалар бетінде орналасқан Н1 және Н2 рецепторлармен әсерлеседі. Гистолиннің негізгі патогенетикалық әсері тегіс бұлшықеттің кенет қысқарылуымен көрсетілінеді. Осы сияқты жиырылу астма немесе анафилактикалық шок кезіндегі бронхотүйілуіне жауапты болады. Гистоминнің тамыр жүйесіне әсер етуі - тамыр қабырғасын жалаңаштау және олардың өткізгіштігін жоғарлату. Басқа медиатор – сратоминнің ағзаға патогенетикалық гистаминге ұқсас. Адамда сиратаминнің активтілігі тек тробоциттер мен ащы ішек клеткаларына байланысты ғана байқалады.
Хематоксикалық факторлар, бұлтты клеткалардың гранулаларынан бөлінетін, реакцияның даму ошағына гранулоциттер мен нейтрофильдердің жиналуын қамтамасыз етеді.
Жоғары сезімталдықтың баяу типі.
IV тип- жоғарғы сезімталдықтың баяу типі (ГБТ)- қабынудың антигенспецификалық СДТ4- клеткалар қызметінің нәтижесі болып табылады. Реакцияның дамуына әкелетін келесі сатыларды ажыратуға болады:
1. антигеннің ағзаға бірінші енуі спецификалық СДТ4- қабыну клеткаларының (Тн1) жинауына әкеледі.
2. антиген қайталап енуінде, оны жергілікті орналасқан тінді макрофагтар жұтады. Осы антигентаныстырушы клеткалар антигенді фрагметтерін ІІ класс МНС молекулалармен комплексте өз беткейіне шығарады.
3. ангтигенспецификалық Тн1- клеткалар макрофог беткейінде иммуногенді комплекспен әсерлеседі. Өткен әсерлесуден кейін қабынудың Т- клеткалары цитокиндер жинағының секрециясын бастайды: макрофагтар миграциясын тежеуші фактор (макрофагингиберлеуші фактор- МИФ), макрофагильді хемотоксикалық фактор (МХФ), 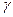 - интерферон және
- интерферон және  - интерферон (ИНФ- және ИНФ- ), интерлейкин- 3 (ИЛ-3) және гарунлоцитарлы- макрофаголды колония стимульдеуші фактор (ГМ- КСФ).
- интерферон (ИНФ- және ИНФ- ), интерлейкин- 3 (ИЛ-3) және гарунлоцитарлы- макрофаголды колония стимульдеуші фактор (ГМ- КСФ).
4. Секреттелуші цитокиндер қабыну реакциясын, оның визуальді көріністерін қамтамасыз етеді. МИФ және МХФ антиген енген аймағына қосымша фагоциттеруші клеткаларды жақындатады. - ИФН қабыну медиаторларының өнімін күшейтуші макрофагтарды белсендіреді. ИНФ- локальді тіндік зақымдануды орнын анықтайды және қабыну аймағындағы қан тамырларындағы адгезивті молекулалардың экспрессиясын күшейтеді, сонысымен қосымша қабыну клеткаларының жеңіл түрде енуіне әсер етеді. ИЛ-3 және ГМ- КСФ гемопоэтикалық дифференцировка факторлары ретінде сүйек кемігі прекурсорынан монциттердің пісіп- жетілуін қамтамасыз етеді. Қайта түзілетін моноцит- макрофагтар, қабыну аймағына миграциялап, өз функциясын атқарып болған соң бұзылатын макрофагтардың санын толтырады. Бұл процестердің барлығы, патогенді оңашалауға бағытталған (немесе қандай да бір басқа антигенді), 24-48 сағатта қабыну ошағы түзілуімен аяқталады.
| <== попередня сторінка | | | наступна сторінка ==> |
| Интерлейкин-12 | | | Аутоиммунитет |
|
Не знайшли потрібну інформацію? Скористайтесь пошуком google: |
© studopedia.com.ua При використанні або копіюванні матеріалів пряме посилання на сайт обов'язкове. |