РЕЗОЛЮЦІЯ: Громадського обговорення навчальної програми статевого виховання
ЧОМУ ФОНД ОЛЕНИ ПІНЧУК І МОЗ УКРАЇНИ ПРОПАГУЮТЬ "СЕКСУАЛЬНІ УРОКИ"
ЕКЗИСТЕНЦІЙНО-ПСИХОЛОГІЧНІ ОСНОВИ ПОРУШЕННЯ СТАТЕВОЇ ІДЕНТИЧНОСТІ ПІДЛІТКІВ
Батьківський, громадянський рух в Україні закликає МОН зупинити тотальну сексуалізацію дітей і підлітків
Відкрите звернення Міністру освіти й науки України - Гриневич Лілії Михайлівні
Представництво українського жіноцтва в ООН: низький рівень культури спілкування в соціальних мережах
Гендерна антидискримінаційна експертиза може зробити нас моральними рабами
ЛІВИЙ МАРКСИЗМ У НОВИХ ПІДРУЧНИКАХ ДЛЯ ШКОЛЯРІВ
ВІДКРИТА ЗАЯВА на підтримку позиції Ганни Турчинової та права кожної людини на свободу думки, світогляду та вираження поглядів
- Гідрологія і Гідрометрія
- Господарське право
- Економіка будівництва
- Економіка природокористування
- Економічна теорія
- Земельне право
- Історія України
- Кримінально виконавче право
- Медична радіологія
- Методи аналізу
- Міжнародне приватне право
- Міжнародний маркетинг
- Основи екології
- Предмет Політологія
- Соціальне страхування
- Технічні засоби організації дорожнього руху
- Товарознавство продовольчих товарів
Тлумачний словник
Авто
Автоматизація
Архітектура
Астрономія
Аудит
Біологія
Будівництво
Бухгалтерія
Винахідництво
Виробництво
Військова справа
Генетика
Географія
Геологія
Господарство
Держава
Дім
Екологія
Економетрика
Економіка
Електроніка
Журналістика та ЗМІ
Зв'язок
Іноземні мови
Інформатика
Історія
Комп'ютери
Креслення
Кулінарія
Культура
Лексикологія
Література
Логіка
Маркетинг
Математика
Машинобудування
Медицина
Менеджмент
Метали і Зварювання
Механіка
Мистецтво
Музика
Населення
Освіта
Охорона безпеки життя
Охорона Праці
Педагогіка
Політика
Право
Програмування
Промисловість
Психологія
Радіо
Регилия
Соціологія
Спорт
Стандартизація
Технології
Торгівля
Туризм
Фізика
Фізіологія
Філософія
Фінанси
Хімія
Юриспунденкция
Рухові рефлекси заднього мозку, децеребраційна ригідність.
Задній мозок – це довгастий мозок та міст. Вони зберігають ознаки сегментарної будови, але мають і надсегментарні утворення. Серед найбільш важливих рухових надсегментарних утворень – присінкові ядра та сітчаста речовина. Вони носять назву надсегментарних утворень, так як впливають на м’язи не прямо, а через мотонейрони сегментарних структур – рухові ядра спинного мозку і черепно-мозкових нервів.
Задній мозок отримує і переробляє всю аферентну інформацію, що надходить від спинного мозку, оскільки всі специфічні висхідні шляхи від спинного мозку, входячи в стовбур мозку (задній та середній мозок) віддають коллатералі (гілочки) до ретикулярної формації Ù тут продовжується обробка аферентної інформації.
Задній мозок отримує нову аферентну інформацію, яка відіграє роль в регуляції рухових функцій – це інформація від вестибулярних рецепторів.

|
 |  | ||||
| |||||


Отолітові рецептори – являють собою волоскові клітини, волоски яких занурені в отолітову мембрану – желеподібна мембрана з включенням кристалів CaCО3 – отолітів. Зміщення цієї мембрани деформує волоски Ù збудження рецептора (чи його гальмування, в залежності від того, в який бік згинається волосок). Отолітова мембрана зміщується відносно волоскових клітин при двох умовах:
1. Зміщення голови в просторі. 2. Рух з лінійним прискоренням.
Вестибулорецептори півколових канальців – волоскові клітини, які локалізуються в ампулах півколових канальців і занурені в ендолімфу цих канальців. Ендолімфа зміщується відносно волоскових клітин і деформує їх при русі з кутовим прискоренням. При цьому збуджуються волоскові клітини канала, який знаходиться в тій площині, в якій проходять обертальні рухи (у відповідності з трьохвимірним простором, в якому ми існуємо, наявні 3 півколових канальці).
Від вестибулорецепторів інформація передається в задній мозок по аферентним волокнам VIII пари черепно-мозкових нервів (далі ЧМН) – вестибуло-кохлеарного нерва (точніше – по його вестибулярній частині). В задньому мозку розміщені 4 вестибулярні ядра (медіальне, латеральне, верхнє та нижнє). В регуляції рухових функцій у людини найбільше значення має латеральне вестибулярне ядро Дейтерса, від якого починається вестибулоспінальний шлях (частина МНС).
Вивчити роль заднього мозку в регуляції рухових функцій можна на тваринах з децеребрацією – перерізкою стовбура мозку між середнім та заднім мозком (точніше – нижче червоних ядер середнього мозку). При порівнянні можливостей регуляції рухових функцій такої тварини і спінальної тварини, можна оцінити роль в регуляції цих функцій саме заднього мозку.
 Після операції децеребрації у тварини виникає децеребраційна ригідність, яка у кішки проявляється у слідуючому: передні і задні кінцівки розігнуті; голова закинута назад за рахунок розгинання шиї; хвіст піднятий; тулуб розігнутий. Ця поза кішки пов’язана з гіпертонусом розгиначів. Позу децеребраційної тварини (децеребраційна ригідність) можна охарактеризувати як посилену позу антигравітації. Згадаємо, що спинний мозок забезпечував формування м’язового тонусу рефлекторної природи, але не забезпечував формування пози антигравітації із-за малої сили тонусу.
Після операції децеребрації у тварини виникає децеребраційна ригідність, яка у кішки проявляється у слідуючому: передні і задні кінцівки розігнуті; голова закинута назад за рахунок розгинання шиї; хвіст піднятий; тулуб розігнутий. Ця поза кішки пов’язана з гіпертонусом розгиначів. Позу децеребраційної тварини (децеребраційна ригідність) можна охарактеризувати як посилену позу антигравітації. Згадаємо, що спинний мозок забезпечував формування м’язового тонусу рефлекторної природи, але не забезпечував формування пози антигравітації із-за малої сили тонусу.
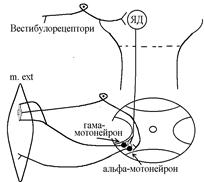 В основі формування децеребраційної регідності лежать вестибулоспінальні впливи. Присінкові ядра Дейтерса тонічно активні, так як вестибулорецептори адаптуються дуже повільно. Від цих ядер по присінково-спинномозковим шляхам, що не перехрещуються, інформація поступає до мотонейронів, які інервують м’язи тулуба та проксимальних відділів кінцівок. При цьому a- та g-мотонейрони екстензорів активуються, а a- та g-мотонейрони флексорів, навпаки – гальмуються.
В основі формування децеребраційної регідності лежать вестибулоспінальні впливи. Присінкові ядра Дейтерса тонічно активні, так як вестибулорецептори адаптуються дуже повільно. Від цих ядер по присінково-спинномозковим шляхам, що не перехрещуються, інформація поступає до мотонейронів, які інервують м’язи тулуба та проксимальних відділів кінцівок. При цьому a- та g-мотонейрони екстензорів активуються, а a- та g-мотонейрони флексорів, навпаки – гальмуються.
Активація a-мотонейронів екстензорів приводить до безпосередньої активації екстрафузальних волокон цих м’язів Ù збільшення їх тонуса. Активація g-мотонейронів приводить до скорочення скоротливих сегментів інтрафузальних волокон Ù скорочення цих сегментів Ù розтягнення ядерної сумки інтрафузального волокна Ù збудження рецептора Ù передача інформації по g-петлі до a-мотонейронів Ù посилення їх активності Ù збільшення тонусу м’язів екстензорів. Протилежні зміни виникають у флексорній групі м’язів.
Тому коротко можна сказати, що в основі розвитку децеребраційної ригідності лежать вестибулоспінальні впливи, завдяки яким відбувається:
- a- та g-посилення міотатичних тонічних рефлексів екстензорів;
- a- та g-послаблення міотатичних тонічних рефлексів флексорів.
Шийні рефлекси положення – виникають у відповідь на зміну нормального положення голови відносно тулуба. При цьому подразнюються пропріорецептори шиї Ù передача інформації в шийні сегменти спинного мозку Ù по пропріоспінальним низхідним шляхам до мотонейронів спинного мозку Ù перерозподіл м’язового тонусу, який змінює положення тулуба відповідно до положення голови:
- якщо голова нахиляється чи повертається в бік, то зі сторони нахилу збільшується тонус розгиначів, а з протилежної сторони – тонус згиначів. Такий перерозподіл тонуса попереджає падіння, тобто втрату пози;
- якщо голова нахиляється вперед, тонус згиначів збільшується на передніх кінцівках кішки, тонус розгиначів – на задніх (кицька п’є молоко чи жере Віскас);
- якщо голова закидається назад, тонус розгиначів збільшується на передніх кінцівках кішки, тонус згиначів – на задніх (кицька дивиться на папугу й готується до стрибка).
Присінкові (вестибулярні) рефлекси положення – виникають при зміні положення голови в просторі. При нахилах чи поворотах голови збуджуються отолітові вестиблорецептори Ù інформація передається до вестибулярних ядер Дейтерса Ù до мотонейронів спинного мозку та до м’язів. В результаті зі сторони нахилу (поворота) збільшується тонус розгиначів, а з протилежної сторони – тонус згиначів Ù збереження пози.
 | |||
| |||

 |  | ||||
| |||||
|
|
|
 | |||
| |||
Перерозподіл тонусу за рахунок шийних рефлексів не можна отримати у спінальної жаби через слабкість тонуса, хоча рівень їх замикання – спинний мозок. Задній мозок підсилює ці рефлекси.
Шийні та присінкові рефлекси положення у вигляді перерозподілу тонуса можна отримати у децереброваної тварини. Але на фоні ригідності ступінь цього перерозподілу недостатній для утримання антигравітаційної пози: децеребрована тварина може стояти, як лялька, але ще не може втримувати рівновагу при умовах, що вмикають шийні та присінкові статичні рефлекси положення.
Таким чином, задній мозок:
1. Забезпечує створення посиленої антигравітаційної пози – пози децеребраційної ригідності.
2. Забезпечує здійснення статичних позових рефлексів (шийних та присінкових), що спрямовані на збереження рівноважної пози при зміні положення голови відносно тулуба (шийні) чи при зміні положення голови в просторі (присінкові), але ступінь виразності цих рефлексів на фоні посиленого тонуса недостатній для збереження пози.
3. Не забезпечує збереження пози при русі (немає стато-кінетичних рефлексів), відновлення порушеної пози (немає випрямляючих рефлексів).
4. Фазичні рефлекси на фоні посиленого тонуса не викликаються.
Ретикулярна формація стовбура мозку, характер її висхідних та низхідних впливів.
Сітчаста речовина (ретикулярна формація – РФ) – скупчення нервових клітин, які мають велику кількість дендритів, що галузяться та довгі аксони, які формують низхідні (ретикулоспінальні) та висхідні шляхи сітчастої речовини.
Аферентних шляхів РФ дуже багато, вона отримує інформацію від:
- усіх специфічних висхідних еферентних шляхів, які віддають колатералі до ядер РФ, заходячи в стовбур мозка;
- від іних утворень ЦНС, а саме:
- від кори головного мозку;
- від гіпоталамуса;
- від мозочка;
- від базальних ядер;
- від інших утворень.
Еферентні зв’язки РФ можна поділити на висхідні та низхідні.
Висхідні зв’язки РФ та висхідні впливи:
- через неспецифічні ядра таламуса передає інформацію всім відділам головного мозку та має неспецифічний активізуючий вплив на клітини. Цей вплив забезпечує бодьорий стан організму та визначає рівень активності клітин кори. РФ має окремі структури (гіпногенні зони), котрі зменшують ступінь (рівень) активності клітин кори та зумовлюють стан сну;
- по ретикулогіпоталамічним шляхам передає інформацію від різних рецепторів в гіпоталамус.
Характер низхідних ретикулоспінальних впливів:
- від медіального ядра РФ заднього мозку починається ретикулоспінальний (входить до складу МНС) – специфічно активує мотонейрони флексорів та гальмує мотонейрони екстензорів тулуба та проксимальних відділів кінцівок;
- від інших ядер РФ беруть початок шляхи, які неспецифічно активують чи гальмують спінальні мотонейрони і таким чином визначають ступінь їхньої активності;
- від вегетативних центрів РФ (дихальний, гемодинамічний) по ретикулоспінальним шляхам передається інформація до симпатичних рухових нейронів спинного мозку при регуляції відповідних функцій організму.
Читайте також:
- Безумовні рефлекси новонароджених
- В) критичність, рефлексивність.
- Вимикач обігріву заднього вікна
- Вища нервова діяльність мавп. Основні особливості умовно-рефлекторної діяльності мавп. Рухові навички і інтелект мавп. Абстракція і узагальнення у мавп.
- Гемодинамічний центр. Рефлекторна регуляція тонусу судин. Пресорні і депресорні рефлекси.
- Глава 4. Психологический анализ урока (занятия) как единство проективно-рефлексивных умений педагога
- Дзеркало заднього огляду в салоні
- Заміна ламп освітлення заднього ліхтаря
- Зіничні рефлекси
- Зіничні РЕФЛЕКСИ
- Зовнішнє дзеркало заднього огляду
- На функції головного мозку, які вимагають додаткового підтвердження.
| <== попередня сторінка | | | наступна сторінка ==> |
| Провідникова функція спинного мозку. Залежність спінальних рефлексів від діяльності центрів головного мозку. Спінальний шок. | | | Рухові рефлекси середнього мозку, їх фізіологічне значення. |
|
Не знайшли потрібну інформацію? Скористайтесь пошуком google: |
© studopedia.com.ua При використанні або копіюванні матеріалів пряме посилання на сайт обов'язкове. |