РЕЗОЛЮЦІЯ: Громадського обговорення навчальної програми статевого виховання
ЧОМУ ФОНД ОЛЕНИ ПІНЧУК І МОЗ УКРАЇНИ ПРОПАГУЮТЬ "СЕКСУАЛЬНІ УРОКИ"
ЕКЗИСТЕНЦІЙНО-ПСИХОЛОГІЧНІ ОСНОВИ ПОРУШЕННЯ СТАТЕВОЇ ІДЕНТИЧНОСТІ ПІДЛІТКІВ
Батьківський, громадянський рух в Україні закликає МОН зупинити тотальну сексуалізацію дітей і підлітків
Відкрите звернення Міністру освіти й науки України - Гриневич Лілії Михайлівні
Представництво українського жіноцтва в ООН: низький рівень культури спілкування в соціальних мережах
Гендерна антидискримінаційна експертиза може зробити нас моральними рабами
ЛІВИЙ МАРКСИЗМ У НОВИХ ПІДРУЧНИКАХ ДЛЯ ШКОЛЯРІВ
ВІДКРИТА ЗАЯВА на підтримку позиції Ганни Турчинової та права кожної людини на свободу думки, світогляду та вираження поглядів
- Гідрологія і Гідрометрія
- Господарське право
- Економіка будівництва
- Економіка природокористування
- Економічна теорія
- Земельне право
- Історія України
- Кримінально виконавче право
- Медична радіологія
- Методи аналізу
- Міжнародне приватне право
- Міжнародний маркетинг
- Основи екології
- Предмет Політологія
- Соціальне страхування
- Технічні засоби організації дорожнього руху
- Товарознавство продовольчих товарів
Тлумачний словник
Авто
Автоматизація
Архітектура
Астрономія
Аудит
Біологія
Будівництво
Бухгалтерія
Винахідництво
Виробництво
Військова справа
Генетика
Географія
Геологія
Господарство
Держава
Дім
Екологія
Економетрика
Економіка
Електроніка
Журналістика та ЗМІ
Зв'язок
Іноземні мови
Інформатика
Історія
Комп'ютери
Креслення
Кулінарія
Культура
Лексикологія
Література
Логіка
Маркетинг
Математика
Машинобудування
Медицина
Менеджмент
Метали і Зварювання
Механіка
Мистецтво
Музика
Населення
Освіта
Охорона безпеки життя
Охорона Праці
Педагогіка
Політика
Право
Програмування
Промисловість
Психологія
Радіо
Регилия
Соціологія
Спорт
Стандартизація
Технології
Торгівля
Туризм
Фізика
Фізіологія
Філософія
Фінанси
Хімія
Юриспунденкция
Будова та функції ДНК, РНК, АТФ.
До полінуклеїнових кислот відносяться дезоксирибонуклеїнова (ДНК) і рибонуклеїнова (РНК) кислоти. Це найважливіші біополімери, які відіграють головну роль у зберіганні, передачі і реалізації генетичної інформації у живих клітинах.
У середині ХІХ століття було встановлено, що матеріал, який відповідає за передачу спадкових ознак, знаходиться у ядрі. А у 1869 р. швейцарський лікар Ф. Мішер (1844 – 1895), працюючи з ядрами клітин гною, виділив речовину, що мала кислі властивості. Цю подію оцінюють як відкриття нуклеїнових (“ядерні”) кислот. Сам термін з’явився дещо пізніше – у 1889 р.
У 1891 р. німецький біохімік А. Кьоссель (1853 – 1927) провів гідроліз нуклеїнових кислот і встановив, що вони складаються з залишку цукру, фосфорної кислоти і азотистих основ, які належать до пуринів і піримідинів. Полімерна природа нуклеїнових кислот була виявлена у 30-х рр. ХХ століття Але знадобилося більше 20-ти років для того, щоб з’ясувати всі ті складні закономірності, які лежать у основі побудови нуклеїнових полімерів. Завдяки дослідженням будови і функціонування цих полімерів у середині ХХ століття виникла нова біологічна наука – молекулярна біологія. Саме з нею пов’язані найвизначніші досягнення біології другої половини ХХ століття.
Полінуклеїнові кислоти належать до складних гетерополімерів, мономерами яких є нуклеотиди. Мономери не лише відрізняються між собою, але і кожен мономер має складну будову.
Значний вклад у розшифрування будови нуклеїнових кислот вніс американський біохімік Е. Чаргафф, який народився у 1905 році у м. Чернівці. Аналізуючи у 50-х рр. ХХ століття склад нуклеїнових кислот різної природи, Е. Чаргафф вивів що:
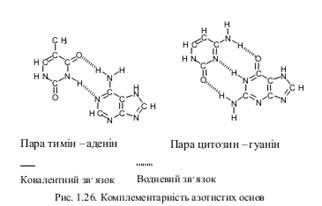
Співвідношення суми аденінів з тимінами до суми гуанінів з цитозинами однакове у ДНК з різних клітин. Ця закономірність відома під назвою правила Чаргаффа. Вона лежить у основі побудови вторинної структури нуклеїнових кислот. Вторинна структура створюється завдяки специфічним водневим зв’язкам, як виникають між азотистими основами.
У зв’язку з тим, що між тиміном (урацилом) виникають два водневі зв’язки, а між цитозином і гуаніном – три, співвідношення між цими парами основ завжди стале.
Пари азотистих основ, між якими виникають водневі зв’язки, називають комплементарними (відповідними), а закономірність їх утворення – принципом комплементарності (відповідності). Таким чином, у випадку, коли виникає вторинна структура нуклеїнових кислот, навпроти тиміну (урацилу) завжди знаходиться аденін, а навпроти цитозину – гуанін.
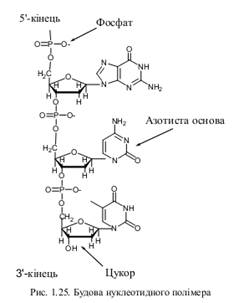
Залежно від природи цукру, нуклеїнові кислоти поділяються на два типи:
рибонуклеїнові (РНК), у складі нуклеотидів яких знаходиться цукор рибоза;
дезоксирибонуклеїнові (ДНК), у складі нуклеотидів яких знаходиться цукор дезоксирибоза.
У побудові цих полімерів існує декілька принципових відмінностей.
На рівні первинної структури полінуклеїнові кислоти відрізняються не лише видом цукру (рибоза чи дезоксирибоза), але і типом азотистих основ. Аденін, гуанін і цитозин входять до складу нуклеотидів як ДНК, так і РНК, але тимін зустрічається лише у ДНК, а урацил – тільки у РНК.
Розміри нуклеїнових кислот залежать від їх типу і функції. Їх визначають трьома різними способами:
через молекулярну масу (у дальтонах);
через кількість нуклеотидів (для РНК – у нуклеотидах, а для ДНК – у парах нуклеотидів);
через константу седиментації (у свебергах, 1 S = 10-13 сек.).
Константа (коефіцієнт) седиментації біологічних макромолекул визначається за допомогою високошвидкісного центрифугування (ультрацентрифугування). Залежно від маси і просторової організації макромолекули осаджуються у центрифужних пробірках з різною швидкістю. Константу седиментації біологічних макромолекул виражають у одиницях Свеберга. Це величина постійна для даного виду макромолекул.
Будова молекули ДНК. Макромолекула ДНК – це два довгі полімерні ланцюги, що складаються з мономерів дезоксирибонуклеотидів, тісно з'єднаних між собою. Нитки ДНК з'єднуються водневими зв'язками між азотистими основами двох ланцюгів і утворюють подвійну спіраль ДНК. Проаналізувавши попередні результати, американець Дж. Уотсон (н. 1928 р.) і англієць Ф. Крік (н. 1916 р.) у 1953 р. запропонували модель тримірної структури ДНК. Це повідомлення молодих дослідників (Ф. Кріку було 35 років, а Дж. Уотсону – лише 25 років) було опубліковане у журналі “Nature”. Воно відіграло важливу роль у розвитку молекулярної біології та генетики. У 1962 р. Дж. Уотсон і Ф. Крік разом з М. Уілкінсом були нагороджені за цю роботу Нобелівською премією у галузі фізіології і медицини.
Характерні особливості просторової організації молекули ДНК («подвійної спіралі») такі:
· Пуринові та піримідинові основи розташовані всередині спіралі й з’єднані між собою водневими зв’язками відповідно принципу комплементарності.
· Залишки фосфату і дезоксирибози з’єднані ковалентними зв’язками і знаходяться назовні від осі спіралі.
· Площина азотистих основ майже перпендикулярна осі спіралі й ланцюгу з фосфатів і цукру.
· Діаметр спіралі 2,0 нм. Відстань між сусідніми основами вздовж осі – 0,34 нм. Таким чином, на один виток спіралі припадає 10 нуклеотидів, що відповідає 3,4 нм.
· Два полінуклеотидні ланцюги утримуються разом завдяки водневим зв’язкам між азотистими основами відповідно до принципу комплементарності.
· Послідовність нуклеотидів у одному ланцюзі не має ніяких обмежень.
· Відповідно до принципу комплементарності, ланцюги ДНК направлені у протилежну сторону (антипаралельні).
Молекула ДНК – місце збереження у клітині спадкової інформації.
Це дволанцюговий лінійний полімер. Вона надзвичайно довга. Якщо витягнути у одну лінію всі ДНК, які містяться у організмі людини, ми отримаємо нитку, яка у 100 разів більша, ніж відстань від Землі до Сонця. Коли означити кожен нуклеотид всіх 46 ДНК клітин людини однією буквою, а потім видрукувати цей текст, то ми отримаємо книжку з більш ніж 820 тисячами сторінок. Надзвичайно великі розміри подвійної спіралі ДНК не дають можливість їй вільно розташовуватися у клітині. Тому для цієї молекули характерне також специфічне упаковування за допомогою білків.
Процес подвоєння ДНК – реплікація (replication) – забезпечує відтворення спадкової інформації та передачу її до дочірніх клітин при мітозі й мейозі. Синтез ДНК відбувається при реплікації з використанням обох полінуклеотидних ланцюгів як матриць – за так званим напівконсервативним механізмом: дві дочірні молекули-копії містять один материнський ланцюг (що слугував матрицею) і один ланцюг, синтезований de novo. Включення нуклеотидів до ланцюга, що синтезується, детермінується матрицею за принципом комплементарності. Такий механізм реплікації став зрозумілим відразу після того, як Уотсоном і Кріком була сформульована модель подвійної спіралі ДНК.
Зростання ланцюга ДНК відбувається в напрямку від 5' до 3'-кінця. Субстратами реакції є 3'- кінцева ОН- група дезоксирибози зростаючого ланцюга та дезоксирибонуклеозидтрифосфати (dNTP). Фермент, що каталізує цю реакцію – ДНК-залежна ДНК-полімераза (DNA dependent DNA Polymerase, DNAP). Синтез ДНК за подібними механізмами здійснюється також при репарації пошкоджень і деяких інших процесах .
Репарація (repair) ДНК – один із загальних біологічних процесів, який спрямований на виправлення помилок синтезу ДНК при реплікації, а також численних пошкоджень, котрі виникають у ДНК унаслідок дії хімічних і фізичних факторів. До таких пошкоджень відносять різноманітні хімічні модифікації азотистих основ, ковалентні зшивки сусідніх піримідинів (утворення піримідинових, найчастіше тимінових, димерів) під дією ультрафіолетового випромінювання, одно- і дволоанцюгові розриви під дією іонізуючої радіації та вільних радикалів тощо. Часто системи репарації працюють під час або відразу після реплікації. Більшість репараційних процесів передбачає видалення пошкодженої одноланцюгової ділянки з наступним синтезом ДНК за допомогою ДНК-полімераз. Але існують і процеси, пов'язані з безпосереднім ì виправленням пошкодженого елемента за рахунок прямої дії певних ферментів.
Суттєвим моментом існування ДНК у живих системах є не тільки процеси відновлення та збереження інформації, що міститься в послідовності нуклеотидів, а й різноманітні операції, спрямовані на перетасування цієї інформації з метою створення нових комбінацій генів. Серед процесів рекомбінації (recombination), які приводять до генетичного розмаїття, розрізняють такі:
• Гомологічна рекомбінація – обмін ділянками між досить довгими молекулами ДНК із гомологічними послідовностями пар основ. Відбувається в усіх організмів, які розмножуються статевим шляхом, між гомологічними хромосомами при мейозі (кросинговер), а також при мітозі та в прокаріотів – наприклад, після кон'югації двох бактеріальних клітин і проникнення ДНК з однієї в іншу .
• Сайт-специфічна рекомбінація – вирізання / вбудовування однієї молекули ДНК із / в іншу, або зміна орієнтації (інверсія) фрагмента ДНК у межах однієї молекули. Рекомбінація цього типу здійснюється за рахунок упізнання специфічними білками коротких елементів послідовності ДНК.
• Незаконна рекомбінація - об'єднання двох молекул ДНК, які не мають ані гомології, ані специфічних елементів послідовності.
• Переміщення в межах геному мобільних елементів послідовності ДНК.
Рибонуклеїнові кислоти (РНК). Спадкова інформація зберігається в молекулі ДНК. Проте ДНК не бере участі в життєдіяльності клітин. Роль посередників у передачі спадкової інформації від ДНК у цитоплазму відіграють рибонуклеїнові кислоти. Взаємовідносини ДНК, РНК і білків можна представити у вигляді схеми ДНК —> РНК —> білок.
Рибонуклеїнові кислоти у клітині виконують функції, пов’язані з копіюванням і реалізацією генетичної інформації. На частку РНК у організмі хребетних припадає 5–10 % від загальної маси нуклеїнових кислот. Найпоширенішими видами РНК є:
інформаційна або матрична (мРНК) – включає фрагмент генетичної інформації і транспортує її з ядра у цитоплазму, бере участь у біосинтезі білка;
рибосомальна (рРНК) – входить до складу рибосом;
транспортна (тРНК) – бере участь у біосинтезі білка.
Вміст РНК у клітині і співвідношення між її окремими видами залежить від інтенсивності поділу і швидкості біосинтезу білка. У клітин з високою метаболічною активністю він вищий, ніж у клітин, які не мають таких властивостей . Але коли клітини активно функціонують, то співвідношення між різними видами РНК визначається їх роллю у метаболічних процесах і приблизно становить:
мРНК – 2–5 %;
рРНК – 80–85 %;
тРНК – 14–16 %.
Молекула РНК, за винятком деяких вірусів, одноланцюгова. Незважаючи на це, вона здатна набувати у просторі певної стабільної форми, яка пов’язана з її функціональною активністю. Через одноланцюговість нуклеотидний склад РНК не підкоряється правилу комплементарності. Але деякі ділянки містять комплементарні послідовності, які здатні до взаємодії. Між ними виникають водневі зв’язки, які приводять до утворення “шпильок”.
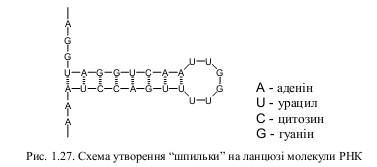
“Шпильки” – це петлі, які утворюються на одноланцюговій молекулі РНК між сусідніми комплементарними ділянками нуклеотидної послідовності. Утворення “шпильок” має велике значення у стабілізації просторової структури РНК. Частка подвоєних ділянок у різних видів РНК змінюється у широких межах, становлячи, у середньому, 50 %.
Серед полімерних рибонуклеїнових кислот найхарактернішу просторову структуру має тРНК. Ця сполука відповідає за доставку і розпізнавання амінокислотних залишків у процесі біосинтезу білка. У 1965 році Р. Холлі встановив первинну структуру однієї з тРНК і запропонував модель її вторинної структури. Вона отримала назву “листок конюшини”.
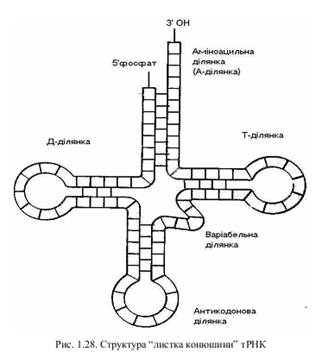
На сьогодні визначено будову більше 100 тРНК, але всі вони виявилися досить подібними. У структурі тРНК одноланцюгові ділянки чергуються дволанцюговими, стабілізованими водневими зв’язками між комплементарними азотистими основами. До 3'–кінцевої ділянки через групу ОН приєднується амінокислота, тому цю гілку називають аміноацильною ділянкою (А-ділянкою). Протилежна до неї гілка несе у собі антикодон, який, відповідно до генетичного коду, визначає тип тРНК. Це антикодонова ділянка. Т-ділянка названа по типовому тиміновому нуклеотиду. Д-ділянка містить два (ди-) модифікованих урацилових нуклеотиди. Невеличка варіабельна ділянка характеризується надзвичайно нестійким складом. Більш детально про функціональну активність тРНК ми поговоримо тоді, коли будемо розглядати процес біосинтезу білка.
Модель типу “листок конюшини” прояснила багато властивостей тРНК, але гідродинамічні характеристики молекули свідчили про те, що вона упакована більш компактно. За допомогою методів рентген-структурного аналізу американські дослідники під керівництвом А. Річа і англійські під керівництвом лауреата Нобелівської премії А. Клуга встановили третинну структуру тРНК. Виявилося, що у просторі “листок конюшини” згинається, нагадуючи латинську літеру L.
Серед різноманітних рибонуклеїнових полімерів, конформаційна організація тРНК досліджена найкраще. Але вчені вважають, що загальні закономірності, знайдені у будові цієї молекули, реалізуються і у інших одноланцюгових полінуклеотидах.
Матрична (інформаційна) РНК – це комплементарні копії фрагментів ДНК, які відповідають працюючим у даний момент генам. Ці лінійні молекули надзвичайно гетерогенні. Вони мають різну молекулярну масу і довжину. У одній еукаріотичній клітині одночасно може знаходитися більше 10 4 різних молекул мРНК.
Рибосомальні РНК – основний компонент рибосом. Вони становлять до 65 % їх маси. Це нуклеопротеїди, які здатні до самозбирання.
Розміри РНК, які входять до складу рибосом, фіксовані і специфічні для прокаріотів і еукаріотів. У прокаріотичних клітинах міститься три види рРНК, а у еукаріотичних – чотири.
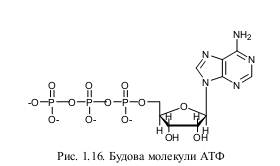
Чи не найвідомішим нуклеотидом є аденозинтрифосфорна кислота (АТФ). Цей нуклеотид складається з рибози, аденіну і трьох фосфатів. АТФ вперше була виявлена в екстрактах м’язових тканин німецьким дослідником К. Ломаном. Майже одночасно, у 1929 р. американські дослідники С. Фіске і Й. Суббароу виділили цю сполуку.
Спочатку вважали, що АТФ приймає участь лише у м’язових скороченнях, але потім ця молекула була виявлена і в інших біологічних об’єктах. У 1941 р. Ф. Ліпман висунув гіпотезу, за якою АТФ в клітинах відіграє роль головного і універсального перетворювача енергії.
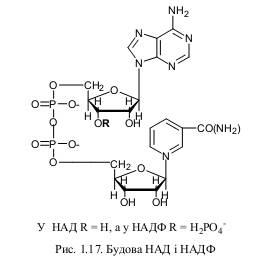
Подальші дослідження підтвердили це блискуче припущення. Крім енергетичної функції, АТФ у клітинах є також універсальним джерелом фосфатних груп, забезпечує транспорт іонів Н+ і деяких інших катіонів через мембрани проти градієнту концентрації тощо. У випадку приєднання до залишку фосфорної кислоти активних груп (наприклад, вітамінів), утворюються інші більш складні нуклеотиди. Всі вони відіграють значну роль у обміні речовин у клітинах. Прикладом такої є нікотинамідаденіндинуклеотид та нікотинамідаденіндинуклеотидфосфат (відповідно, НАД і НАДФ). Вони, подібно до АТФ, приймають участь у енергетичних процесах. НАД і НАДФ складаються з аденіну, двох молекул рибози, з’єднаних двома фосфатними залишками і вітаміну В5. У такому вигляді ці сполуки є активаторами (коферментами) майже 150 біохімічних реакцій, пов’язаних з синтезом енергії та іншими важливими перетвореннями у клітинах як прокаріотів, так і еукаріотів. Відкрили НАД і НАДФ німецькі дослідники Р. Кун і О. Варбург. Крім перерахованих, у клітині є й інші енергетичні нуклеотиди. За своєю будовою вони подібні або до АТФ, або до НАДФ. Таким чином, нуклеотиди є універсальними компонентами обміну речовин і перетворення енергії у біологічних об’єктах.
| <== попередня сторінка | | | наступна сторінка ==> |
| Досягнення сучасної біології. Методи біологічних досліджень. | | | Основні відомості про пасивні кола. |
|
Не знайшли потрібну інформацію? Скористайтесь пошуком google: |
© studopedia.com.ua При використанні або копіюванні матеріалів пряме посилання на сайт обов'язкове. |