РЕЗОЛЮЦІЯ: Громадського обговорення навчальної програми статевого виховання
ЧОМУ ФОНД ОЛЕНИ ПІНЧУК І МОЗ УКРАЇНИ ПРОПАГУЮТЬ "СЕКСУАЛЬНІ УРОКИ"
ЕКЗИСТЕНЦІЙНО-ПСИХОЛОГІЧНІ ОСНОВИ ПОРУШЕННЯ СТАТЕВОЇ ІДЕНТИЧНОСТІ ПІДЛІТКІВ
Батьківський, громадянський рух в Україні закликає МОН зупинити тотальну сексуалізацію дітей і підлітків
Відкрите звернення Міністру освіти й науки України - Гриневич Лілії Михайлівні
Представництво українського жіноцтва в ООН: низький рівень культури спілкування в соціальних мережах
Гендерна антидискримінаційна експертиза може зробити нас моральними рабами
ЛІВИЙ МАРКСИЗМ У НОВИХ ПІДРУЧНИКАХ ДЛЯ ШКОЛЯРІВ
ВІДКРИТА ЗАЯВА на підтримку позиції Ганни Турчинової та права кожної людини на свободу думки, світогляду та вираження поглядів
- Гідрологія і Гідрометрія
- Господарське право
- Економіка будівництва
- Економіка природокористування
- Економічна теорія
- Земельне право
- Історія України
- Кримінально виконавче право
- Медична радіологія
- Методи аналізу
- Міжнародне приватне право
- Міжнародний маркетинг
- Основи екології
- Предмет Політологія
- Соціальне страхування
- Технічні засоби організації дорожнього руху
- Товарознавство продовольчих товарів
Тлумачний словник
Авто
Автоматизація
Архітектура
Астрономія
Аудит
Біологія
Будівництво
Бухгалтерія
Винахідництво
Виробництво
Військова справа
Генетика
Географія
Геологія
Господарство
Держава
Дім
Екологія
Економетрика
Економіка
Електроніка
Журналістика та ЗМІ
Зв'язок
Іноземні мови
Інформатика
Історія
Комп'ютери
Креслення
Кулінарія
Культура
Лексикологія
Література
Логіка
Маркетинг
Математика
Машинобудування
Медицина
Менеджмент
Метали і Зварювання
Механіка
Мистецтво
Музика
Населення
Освіта
Охорона безпеки життя
Охорона Праці
Педагогіка
Політика
Право
Програмування
Промисловість
Психологія
Радіо
Регилия
Соціологія
Спорт
Стандартизація
Технології
Торгівля
Туризм
Фізика
Фізіологія
Філософія
Фінанси
Хімія
Юриспунденкция
Тема 6.
Тема 5
ВЕРТИКАЛЬНІ СТРУКТУРИ ГЕОСИСТЕМИ:
СКЛАД ТА ДЕКОМПОЗИЦІЯ (топічна ландшафтна екологія)
Основні питання:
1.Основні положення.
2.Основні способи декомпозиції.
Вертикальні межі геосистем.
5.1. Основні положення
Мета:вивчити основні положення геосистеми, поняття вертикальної (топічної) структури, концепцію множинності вертикальних структур, основні терміни.
План:
1.Поняття вертикальної (топічної) структури.
2.Концепція множинності вертикальних структур.
3.Елементи вертикальних структур.
Поняття вертикальної (топічної) структури. При аналізі вертикальної (синонім — топічної) структури геосистеми вважається, що вона однорідна в територіальному відношенні, але «по вертикалі» розкладається на різнорідні частини (рослинність — грунт — гірські породи тощо або різні яруси рослинності — горизонти ґрунту — верстви гірських порід тощо), які пов'язані між собою певними відношеннями. Структури подібного типу називають вертикальними. Під складовими вертикальних структур мають на увазі не стільки різні за своїм висотним положенням шари геосистеми, скільки деякі її частини, специфічні в ній за функцією, фізико-хімічними та іншими характеристиками. Такі різні частини можуть займати в геосистемі спільний «висотний поверх» (як, наприклад, трав'яні рослини та наземні тварини), а деякі — пронизувати весь її вертикальний розріз (гази, волога).Тому термін «вертикальна структура геосистеми» слід вважати дещо умовним.
При аналізі вертикальної структури геосистеми будь-яка з її складових розглядається як територіально однорідна, тобто припускається, що її характеристики на певній площі лишаються незмінними (з математичної точки зору є просторово зосередженими параметрами). Зміна значень цих параметрів визначається взаємодією між елементами вертикального розрізу геосистеми.Насправді ж припущення щодо несуттєвості внутрішньотериторіальних відмінностей може бути справедливим хіба що для геосистеми елементарного рівня — геотопу (фації). Проте це не виключає можливості аналізу вертикальних структур і геосистем вищих таксономічних рангів, якщо необхідно зосередити увагу не на їх внутрішньотериторіальних відмінностях, а на взаємодії компонентів природи або деяких інших частин їх вертикальної будови. Такий підхід, наприклад, використовується при аналізі водного балансу геосистем річкових басейнів, теплового та інших балансів ландшафтних зон тощо.
Концепція множинності вертикальних структур. Щоб виділити вертикальну структуру геосистеми, необхідно визначити множину її елементів і тип відношень (зв'язків) між ними. Внутрішньогеосистемні зв'язки надзвичайно різноманітні. Багато з них зумовлені потоками різних речовин та форм енергії, деякі — фізико-хімічними, біохімічними взаємодіями, інші — відношеннями між популяціями організмів та їх окремими особинами, важливі також генетико-еволюційні зв'язки між різними геокомпонентами та їх частинами тощо. В основі цих відношень лежать різні закономірності і тому аналіз кожного з них пов'язаний із специфічним аспектом розгляду вертикальної будови геосистеми, тобто приводить до виділення власної структури. Це означає, що в одній геосистемі можна виділити кілька вертикальних структур різних типів. Геосистема є поліструктурною у вертикальному відношенні.
Тип внутрішньогеосистемних відношень є основою виділення певної вертикальної структури геосистеми, оскільки визначає: 1) найдоцільніший спосіб її поділу на елементи; 2) характер зв'язків між ними; 3) варіанти групування елементів у більші структурні одиниці — підсистеми, компоненти, модулі (тобто способи декомпозиції вертикальної структури).
Оскільки внутрішньогеосистемних зв'язків дуже багато, то тільки ж має бути виділено й відповідних їм структур. Проте це е виключає можливості визначити деякі загальні типи вертикальних структур геосистеми. Така можливість ґрунтується на близькості багатьох процесів за їх фізичною суттю, характером перебігу, змінами геосистеми, які зумовлені цими процесами. Відповідно й способи структуризації вертикального розрізу геосистеми для таких зв'язків однотипні. Всю множину внутрішньогеосистемних зв'язків умовно можна поділити на такі типи: 1) генетико-еволюційні; 2) зумовлені потоком енергії та її трансформацією; 3) зумовлені речовинними потоками (міграцією речовин); 4) відношення тісного кореляційного або інформаційного зв'язку характеристик геосистеми.
Кожний з цих типів відношень визначає відповідний підхід до виділення вертикальних структур геосистеми (її структуризації). Ці підходи різняться принципом, покладеним в основу виділення структурних частин геосистеми (її елементів, компонентів тощо). Розрізняють три загальних підходи структуризації геосистеми та відповідно три типи її вертикальних структур: 1) геокомпонентний (поділ вертикального розрізу геосистеми за компонентами природи і далі за їх генетично однорідними частинами); 2) речовинно-фазовий (структурні частини виділяються як тіла, однорідні за фазовим станом, фізико-хімічними та іншими властивостями речовини) 3) просторово-об'ємний (вертикальний профіль геосистеми поділяється на деякі однорідні шари, точніше — об'єми).
Аналіз кореляційних та інформаційних зв'язків геосистеми (виконується методами математичної статистики та теорії інформації) також приводить до виділення деяких структур (графічно вони зображаються у вигляді кореляційних плеяд або графів). Проте елементами цих структур є не деякі фізичні тіла (частини) геосистеми, а її окремі характеристики. Відношення фіксують лише наявність між ними певного кореляційного зв'язку (або каналу передачі інформації), фізичний зміст якого може бути невідомим та й взагалі при такому аспекті аналізу геосистеми значення не має. Для побудови такого типу структур необхідна не структуризація геосистеми (її поділ на частини), а обґрунтування певного набору її характеристик (змінних). Врешті для будь-якого з наведених трьох типів вертикальних структур геосистеми. можна оцінити ступінь кореляційного (або інформаційного) зв'язку між характеристиками їх елементів і побудувати статистичний (або інформаційний) варіант цих структур.
Елементи вертикальних структур. Перш ніж виділити елементи вертикальних структур геосистеми, необхідно визначити, який принцип оцінки однорідності прийнято за основу структуризації геосистеми (генетичний, речовинно-фазовий тощо) та який рівень детальності структурного аналізу геосистеми достатній для розв'язання поставленого завдання. Вирішення першого з цих питань визначається тим із загальних типів вертикальної структури геосистеми, який аналізується. Щодо другого питання слід зауважити, що мета будь-якого конкретного дослідження геосистеми завжди безпосередньо або опосередковано визначає деяку граничну межу її структуризації, тобто задає такі об'єкти вертикальної структури геосистеми, аналіз внутрішньої будови яких для даного дослідження вже не має значення.Загальних критеріїв визначення таких об'єктів немає, їх обґрунтування залежить від досвіду фахівців.
Спроби віднайти якісь «об'єктивно існуючі» (незалежні від конкретного дослідження), ніби задані самою геосистемою, її неподільні частини (ландшафтні «атоми», «кристали», «геоелементи» тощо), а також деякі «універсальні» критерії елементарності були як в екології, так і в ландшафтознавстві. Незважаючи на деякі цікаві в плані теорії результати цих спроб, зайвий раз переконуємось, що визначення елементарного об'єкта системи залежить тільки від мети та бажаної детальності аналізу. Ним може бути і рослинність взагалі (при загальнотеоретичному аналізі геосистеми), і певна екологічна група видів рослин, і окремий вид, і, окрема рослина, навіть її окремі морфологічні органи.
5.2.Основні способи декомпозиції
Мета:вивчити основні способи декомпозиції, вертикальні межі геосистем, основні терміни.
План:
1.Основні способи декомпозиції.
2.Вертикальні межі геосистем.
Геокомпонентний спосіб. Традиційним для ландшафтознавства поділом геосистеми (ПТК) на складові частини є виділення в ній компонентів природи, кожний з яких, за висловом А.Г. Ісаченка (1991), «є представником окремих геосфер, що складають географічну оболонку». Це гірські породи (представники літосфери), поверхневі та ґрунтові води (гідросфери), повітряні маси (атмосфери), ґрунти (педосфери), рослинність, тварини, мікроорганізми (представники біосфери). Усі ці компоненти є матеріальними тілами. Крім них, А.Г. Ісаченко та деякі інші географи як компоненти природи розглядають також рельєф і клімат. Є пропозиції компонентами ПТК вважати і сукупність продуктів діяльності людини, тісно пов'язаних з природними елементами (таких, як меліоративні канали, шляхи сполучення тощо). Тобто в ландшафтознавстві чіткої визначеності того, що слід вважати компонентом природи і яке їх число, немає.
Ландшафтно-екологічному підходу щодо цього найбільш відповідають погляди Д.Л. Арманда (1975) та В.Б. Сочави (1979). Геокомпонентами вважають матеріальні тіла природного походження, які відрізняються між собою переважаючим фізико-агрегатним (фазовим) станом речовини, наявністю (або відсутністю) та формою органічного життя, основними механізмами утворення, положенням щодо земної поверхні та основними функціями в геосистемі.
Виходячи з цього, до геокомпонентів не належать рельєф і клімат (оскільки це не матеріальні тіла, а їх властивості, які й враховуються при аналізі геосистем) та антропогенні об'єкти як тіла неприродного походження (враховуються як зовнішній по відношенню до геосистем фактор). Разом з тим поверхневі та ґрунтові води слід розглядати не як один геокомпонент (води), а два різних, оскільки вони суттєво відрізняються за своїми функціями в геосистемі, положенням щодо земної поверхні та механізмом утворення. Як самостійні розглядаються й біотичні компоненти — рослинність, тваринний світ та мікроорганізми.
Таким чином, геокомпонентами є тверді маси земної кори, повітряні маси атмосфери, поверхневі та ґрунтові води, ґрунти, рослинність, тварини, мікроорганізми.
Геокомпоненти — складні тіла. У кожному з них є й речовини, які відіграють функцію основної субстанції інших геокомпонентів. Наприклад, під повітряними масами атмосфери слід розуміти не просто суміш газів, а складну субстанцію, що містить також водяну пару, часточки твердих речовин, мікроорганізми. Ще складніший грунт. Ця особливість геокомпонентів надає їм нових — емерджентних властивостей, яких немає в хімічно чистих та однорідних речовинах, що їх складають. А.Г. Ісаченко (1991) зауважує, що в системі організації речовини Землі геокомпоненти займають. проміжне положення між простими дискретними тілами мінералами, газами, ґрунтовими агрегатами, окремими організмами та ін.) та геосистемами. Тому аналіз вертикальної структури геосистеми, складовими якої є геокомтюненти, буде ефективним при виявленні генетико-еволюційних закономірностей геосистем.
Виділення елементів у геокомпонентній вертикальній структурі виходить з поділу геокомпонентів на їх більш генетично однорідні частини. У гірських породах такими елементами є їх окремі літолого-стратиграфічні верстви (виділяються як породи одного віку та походження); у ґрунті — його генетичні горизонти. Ґрунтові води доцільно розрізняти за шаром порід, які їх вміщують, оскільки їх хімічні та інші характеристики генетично зумовлені властивостями водовміщуючої товщі.
Елементами рослинності та тваринного світу геосистеми зручно вважати окремі ценопопуляції (сукупність особин одного виду в межах елементарної геосистеми). При дуже детальному аналізі вертикальної структури геосистеми цей рівень може виявитися недостатнім. Тоді окремі популяції слід розділити на різні вікові групи і як елементи розглядати сукупність особин одного виду певної вікової групи. Доцільно розглядати як елементи і окремі екотипи рослин — групи особин у межах одного виду, які пристосовані до специфічних умов існування за рахунок спадково закріплених особливостей.При більш загальному аспекті генетико-еволюційного аналізу геосистеми розгляд її окремих ценопопуляцій як елементів вертикальної структури може бути занадто докладним. Як елементи в таких випадках можна вважати сукупності декількох ценопопуляцій, наприклад, видів, які належать до однієї флор-генетичної групи або до екоморфи певного типу (екоморфа — життєва форма рослин, зумовлена факторами зовнішнього середовища; цей термін ввів український геоботанік О.Л. Бельгард, 1950) —ксерофіти, мезофіти, гігрофіти, псамофіти та ін., або ж до певного систематичного таксону — злакові, бобові та ін. Принцип, за яким популяції об'єднуються в групи і далі розглядаються як один елемент, залежить від конкретної мети дослідження.
Поверхневі води та повітряні маси при генетико-еволюційному аналізі геосистем на елементи, як правило, не розкладаються.
Речовинно-фазовий (геомасовий) спосіб. При .аналізі потоків певних речовин, їх взаємопереходів та інших форм взаємодії більш виправдана структуризація геосистеми, яка виходить з того, що вона являє собою складну композицію речовин, різних за фазовим станом, фізичними властивостями, хімічним складом.Взаємодії між ними зумовлюють різні процеси в геосистемі (наприклад, продуційний, засолення ґрунтів тощо) і тому, розглядаючи різні речовини як окремі елементи геосистеми, можна ефективно досліджувати механізми внутрішньогеосистемних зв'язків.
Речовинно-фазовий підхід до структуризації природних систем широко використовується в екології, особливо при імітаційному моделюванні екосистем. У найбільш популярному в екології методі такого моделювання (метод системної динаміки; його розробив Дж. Форрестер) як елементи екосистеми виділяються її окремі речовини, локалізовані в певних фізичних тілах. За термінологією методу системної динаміки, такі елементи називаються резервуарами, й основною їх характеристикою є кількість речовини в резервуарі. Як окремі резервуари виділяються, наприклад, «вода у кореневому шарі ґрунту», «вода у транспортно-скелетних органах рослин», «азот у ґрунті», «азот у трав'яних рослинах» тощо. Резервуари, між якими мають місце потоки певної речовини або хімічного елемента (води, азоту тощо), об'єднуються в більші структурні одиниці екосистеми — блоки, або субсистеми.
Близький підхід до структуризації елементарної геосистеми, обґрунтований Н.Л. Беручашвілі (1980), пов'язаний з виділенням геомас. Під ними розуміють якісно своєрідні тіла геосистеми, які мають певну масу, специфічне функціональне призначення, а також швидкість змін у часі та (або) переміщення в просторі. Як геомаси виділяються аеромаси, гідромаси, педомаси, літомаси, фітомаси, зоомаси, мортмаси (мертва органічна речовина). Від геокомпонентів вони відрізняються більшою речовинною однорідністю. Наприклад, під педомасою розуміють не грунт, а тільки ґрунтовий дрібнозем з гумусом, тобто органо-мінеральну суміш, до якої не входять волога ґрунту, його скелетна частина, перові гази, корені рослин, тваринне населення. До аеромаси відносять сухе повітря — суміш газів без водяної пари та аерозолів. Аеромаси містяться не тільки в атмосфері, а пронизують усі геокомпоненти. Аналогічно й гідромаси, зосереджені не лише в поверхневих та ґрунтових водах, а і в інших геокомпонентах. Мортмаса взагалі не має аналогів серед геокомпонентів і являє собою сукупність накопичених відмерлих решток рослин, тварин, їх екскрементів, мікроорганізмів тощо.
При речовинно-фазовій структуризації геосистеми геомаси слід, розглядати як окремі компоненти її вертикальної будови, оскільки деякі їх частини можуть значно відрізнятися за фізичними, хімічними та іншими показниками. Наприклад, фітомаса представлена такими досить характерними частинами, як зелене листя рослин, корені, транспортно-скелетні органи (стовбури та гілки), генеративні органи, лишайники, мохи, мікроорганізми тощо. Гідромаса також складається з різних мас, що відрізняються насамперед середовищем, де вони розміщені (в атмосфері, ґрунті, ґрунтових водах тощо). Тому при детальному аналізі геосистем геомаси поділяються на елементи залежно від агрегатного складу, функціонального призначення, хімічного складу, положення у вертикальному профілі геосистеми, щільності, метричних та інших особливостей. Ступінь детальності поділу геомас на елементи визначається конкретним завданням ландшафтно-екологічного аналізу.
Крім поділу геомас на елементи, Н.Л. Беручашвілі вважає за важливе також і їх класифікацію. За розробленою ним таксономічною схемою, кожна геомаса поділяється на типи, роди і види. Типи геомас виділяються на основі відмінностей у функціональному призначенні в геосистемі, щільності та швидкості зміни в часі і переміщенні в просторі. Наприклад, серед фітомас виділяються такі їх типи, як однорічне листя деревно-чагарникових рослин, багаторічне листя цих рослин, хвойне листя, транспортно-скелетні органи, корені тощо. При диференціації на типи педомас за основу взято відмінності в механічному складі (глинисті, суглинкові, піщані типи педомас тощо), аеромас — їх температуру (кріотермальні, нанотермальні, мезотермальні та інші аеромаси), гідромас — стан вологи та її знаходження в інших геомасах (атмосферні, снігові, льодові, ґрунтові та інші гідромаси). Літомаси поділяються за їх щільністю та хімічним складом (карбонатні, силікатні та інші типи), мортмаси — за ступенем розкладу та походженням (сухостій, підстилка, торф, мор та ін.).
Роди геомас розрізняються у межах типу переважно за інтенсивністю процесів функціонування. Так, у листяних типах фітомас виділяються їх різні роди за вмістом вологи в листі (гідрофітні, мезофітні, ксерофітні та ін.). Педомаси поділяються на роди за вмістом гумусу (високо-, середньо-, малогумусні) та його характером (кальцієві мюллеві, лісові мюллеві, модерні педомаси). Нарешті, види геомас виділяються з урахуванням метричних характеристик (форми, розмірів, орієнтації тощо) їх елементів.
Просторово-об'ємний (геогоризонтний) спосіб. Дослідження вертикальних потоків енергії та речовин у геосистемі, її динамічних змін протягом року пов'язані з урахуванням просторової неоднорідності геосистеми у вертикальному напрямку — її ярусної будови. Хоч певне уявлення про ярусну будову дає поділ геосистеми на її геокомпоненти, ці структурні одиниці все-таки накладаються одна на одну, до того ж вони неоднорідні по вертикалі. Разом з цим у геосистемі досить чітко виділяються певні її шари, майже однорідні за складом різних геомас, та специфічні в інших відношеннях (зокрема, за умовами життя та екологічними процесами).
Вперше яруси, шари в екосистемі виділив В. Шелфорд (1912), але на основі переважно біотичних критеріїв. З більш комплексних позицій до цього питання підійшли брати К. і В. Арнольді (1963). Ю. П. Бяллович (1947, 1960) назвав такі шари хорогоризонтами, а пізніше - біогеогоризонтами і визначив їх як елементарні — далі неподільні по вертикалі частини біогеоценозу. У ландшафтній екології та геофізиці ландшафту близькі концепції майже одночасно запропонували в Грузії, Франції та Німеччині. Н.Л. Беручашвілі, на початку 70-х років обґрунтував поняття геогоризонту та розробив методику виділення геогоризонтів (1974, 1976). Ж. Рішар (1978) подібні одиниці назвав хоплексолями, а їх характерне поєднання—холоплексіоном. Г. Ноймайстер (1981), аналізуючи взаємодії геотопів поблизу Лейпціга, як елементи їх вертикального профілю виділив однорідні шари, а при дослідженні геотопів як тривимірних об'ємів — однорідні просторові тіла.
Геогоризонт розглядається як комплексне утворення, в яке входять усі геомаси, які містяться в певному шарі геосистеми. Цим вони відрізняються від ярусів фітоценозу, оскільки, крім рослин (фітомаси), включають також повітряні і гідромаси, якщо вони в певний період там є (наприклад, шапки снігу на гілках дерев). Основним критерієм виділення геогоризонту є специфічний набір геомас у межах певного шару геосистеми. Зміна цього набору (поява нового виду геомаси, зміна їх пропорцій тощо) свідчить про появу у вертикальному профілі геосистеми нового геогоризонту.
Суттєвими ознаками при виділенні й характеристиці геогоризонтів Н.Л. Беручашвілі вважає ландшафтно-геофізичні параметри: текстуру, щільність, об'єм, колір, оптичні та ін. З ландшафтно-екологічної точки зору геогоризонти мають бути однорідними і за біофізичними, едафічними, ландшафтно-геохімічними показниками. При виділенні геогоризонтів Н.Л. Беручашвілі ці знаки провідними не називає, хоч за розробленою ним методикою геогоризонти виділяються настільки дрібні, що є всі підстави вважати їх однорідними за широким колом не тільки ландшафтно-геофізичних ознак. Основою для виділення ґрунтових геогоризонтів Н.Л. Беручашвілі бере фізичні показники (механічний склад, щільність), гумусовий стан та насиченість коренями рослин. Проте міграція — акумуляція різних речовин у ґрунті визначається не тільки цими його властивостями, а й ландшафтно-геохімічними умовами різних шарів ґрунтового профілю. Особливе значення мають ландшафтно-геохімічні бар'єри поверхні, що розділяють вертикальний профіль ґрунту на шари, які дуже відрізняються за умовами міграції різних хімічних елементів і сполук (Перельман, 1975, Глазовська, 1978). На ландшафтно-геохімічних бар'єрах інтенсивно накопичуються елементи, рухливість яких при досягненні бар'єру суттєво знижується або стає зовсім неможливою, тобто в сусідній шар ґрунту ці елементи не переходять зовсім або ж мігрують із значно меншою інтенсивністю. Таким чином, крім фізичних характеристик ґрунту при виділенні геогоризонтів слід зважати і на ландшафтно-геохімічні бар'єри.Це важливі межі вертикального профілю геосистеми, що розділяють його на шари з досить різними умовами міграції, акумуляції та взаємодії хімічних речовин.
Кожний геогоризонт стисло описується в індексній формі(мал. )і його можна віднести до певного класу, типу, роду та виду. Ці таксономічні одиниці виділяються за домінуючим (або домінуючими) у даному геогоризонті класом, типом, родом, видом, геомас (наприклад: клас — аерофітогоризонт, тип — нанотермальний транспортно-скелетний і т. д.). За характерним набором типів геогоризонтів класифікуються і вертикальні просторово-об'ємні структури геосистем. Основними їх характеристиками Н.Л. Беручашвілі вважає клас геомас, який визначає головні особливості геосистеми, — потужність (відстань від нижньої до верхньої межі елементарної геосистеми), складність (число геогоризонтів у геосистемі), напруженість (число геогоризонтів на 1 м вертикального профілю).
На відміну від ярусів рослинності, генетичних горизонтів ґрунту, літолого-стратиграфічних шарів, геогоризонти змінюються протягом року. Мінлива не тільки їх потужність, а й кількість (так, узимку щезають геогоризонти, основний об'єм яких займає фітомаса листя). Таким чином, тип вертикальної структури змінюється протягом року, причому можна визначити і дати зміни цих типів (як для конкретного року, так і середньо-багаторічні). На цій підставі ґрунтується ефективний підхід до комплексного аналізу річної динаміки геосистем і виділення станів цієї динаміки.
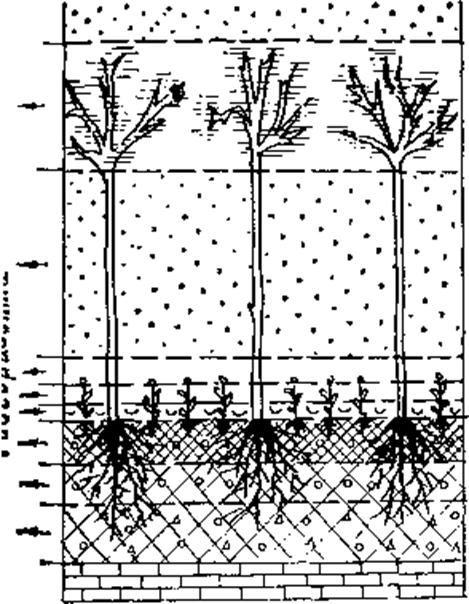
Мал. . Схема виділення геогоризонтів (за Н.Л. Беручашвілі, 1990)
Вертикальні межі геосистем
Основні підходи до виділення меж. Вертикальні межі відокремлюють геосистему від її зовнішнього середовища, точніше — від деяких нижніх верств літосфери (нижня межа) та верхніх шарів атмосфери (верхня межа геосистеми). Вважається, що ці шари мало залежать від внутрішньо-геосистемних процесів. Залежно від аспекту аналізу геосистеми та конкретного завдання дослідження її вертикальні межі можна виділяти за дещо різними критеріями. Якщо геосистема досліджується як певний фізичний об'єм природної реальності, її межами будуть деякі поверхні, при цьому слід чітко визначити їх висоту та глибину щодо земної поверхні. При теоретико-системному аналізі геосистеми, тобто аналізі її деякої схематизованої моделі, склад елементів геосистеми, як правило, задається заздалегідь і таким чином її вертикальні межі визначаються ніби самі собою (як усе те, що не входить до її складу). Аналогічна ситуація виникає й тоді, коли умови деякого конкретного дослідження однозначно визначають склад елементів геосистеми, аналіз яких вважається достатнім для вирішення поставленого завдання. У таких випадках, обґрунтовано визначивши попередньо склад елементів геосистеми, можна правильно виділити і її вертикальні межі. При цьому навіть немає необхідності визначати місцеположення цих меж (тобто їх висоту та глибину). Але це визначення набуває суттєвого значення при фізично-об'ємному аналізі геосистем, особливо при їх стаціонарному дослідженні.
Проте, як місцеположення вертикальних меж залежить від рангу геосистеми, у дослідників визначеності немає. Більшість із них (Н.Л. Беручашвілі, В.О. Боков, К.М. Дьяконов, О.Ю. Ретеюм, В.Б. Сочава, Г. Хаазе та ін.) вважають, що чим вищий ранг геосистеми, тим більша її вертикальна потужність, К.М. Дьяконов (1971) так мотивує цю позицію. Вертикальні межі слід проводити по тому рівню, на якому щезають горизонтальні відмінності між геосистемами. Для фацій лісотундри ці відмінності практично не фіксуються, починаючи з висоти 4—5 м і глибини 2 м. На цих рівнях, на його думку, і слід проводити відповідно верхню та нижню межі фацій. У геосистемах вищого таксономічного рангу — урочищах територіальні відмінності та взаємодії спостерігаються до висоти 7—9 м та глибини 4 м, що й приймається за вертикальні межі цих геосистем. У лісових фаціях О.Ю. Ретеюм (1966) вказує місцеположення їх вертикальних меж на висоті кількох десятків метрів, в урочищах — до кількох сотень, ландшафтах 800—2000 м. Близькі до цих цифри наводить і В.Б. Сочава (1974).
Верхні межі. Характерні особливості верхніх меж геосистем — це їх мінливість у часі залежно від пори року, погодних умов та стану розвитку фітоценозу, а також слабка вираженість цих меж, зумовлена значною відкритістю геосистем у вертикальному напрямку.
При дослідженні зв'язків між елементами геосистеми, зумовлених фізичними процесами (вологообігом, потоками енергії тощо), як складову геосистеми слід розглядати деякий об'єм атмосфери, де ці процеси відбуваються і впливають на його стан. У мікрокліматології такий об'єм атмосфери називають діяльним шаром. Від вищих атмосферних шарів його відрізняють різкі добові коливання метеоелементів, їх неперіодичні зміни, значні вертикальні градієнти, специфічний склад мікрофлори та хімічних елементів повітряних мас, суттєва зміна швидкості та напрямку вітру тощо. Усі ці особливості зумовлені властивостями геосистеми, особливо її альбедо та характером рослинного покриву (його висотою, густиною стояння, іншими фітометричними характеристиками). Фітокліматичними та стаціонарними дослідженнями геосистем встановлено, що їх вплив на значення метеоелементів сягає в 1,5— 2 рази більшої висоти, ніж висота рослинного ярусу.Взимку вплив засніженої поверхні геосистеми на атмосферу набагато менший і загалом не перевищує кількох метрів. Ці орієнтири і можна прийняти при визначенні положення верхньої границі геосистем топічного та хоричного рівнів, проте слід мати на увазі, що це положення мінливе в часі і може змінюватися не тільки протягом року, а й доби.
Верхня межа геосистем регіональної розмірності визначається складніше. Так, досліджуючи ландшафтно-екологічні закономірності процесів переносу та випадання атмосферних забруднень, до складу геосистеми слід включити і той шар тропосфери, в межах якого відбуваються місцеві процеси циркуляції повітря. Висота цього шару визначається не тільки рельєфним фактором, а й станом атмосфери (зокрема, типом циркуляції — циклональним або антициклональним), тому вона також дуже мінлива і може сягати висоти тропопаузи (в середньому 11 км).
При дослідженні біотичних процесів геосистеми, зокрема її продуктивності, за верхню межу можна прийняти межу верхнього рослинного ярусу (аерофітогоризонту), приймаючи шар турбулентної атмосфери безпосередньо над рослинним покривом за зовнішнє середовище. У такий самий спосіб доцільно визначати й верхню межу геосистеми при дослідженні її ґрунтових процесів, зокрема міграції та акумуляції різних речовин у ґрунтовій товщі, водах, рослинах. Роль атмосферних процесів при цьому дуже значна, проте розглядається як фактор зовнішнього середовища.
Нижні межі. При аналізі генетико-еволюційних рис геосистем, які багато в чому визначаються геологічною будовою, у їх нижні межі обов'язково слід включати ті товщі гірських порід, які зумовили становлення даної геосистеми. У цьому плані положення нижньої межі геосистеми залежить від її рангу. Так, геосистеми регіональної та вищої розмірностей можуть бути пов'язані з великими геоструктурами, які своїми коренями заглиблюються в літосферу на десятки кілометрів. При з'ясуванні генезису геосистем хоричної та нижчих розмірностей такі геоструктури виступають у ролі зовнішнього фону, на якому сформувались певні літолого-стратиграфічні та геоморфологічні особливості значно меншої вертикальної потужності. Саме ці особливості й визначили специфіку генезису хоричних і топічних геосистем, для генетико-еволюційного аналізу яких зовсім не обов'язкове занурення в глибини літосфери.
Нижню межу геосистем при їх генетико-еволюційному аналізі здебільшого проводять по гірських породах, які є субстратом формування сучасного рельєфу. В Україні, наприклад, це породи, які залягають під лесовою товщею (вапняки, середньо-верхньо-пліоценові глини тощо), або ж сама ця товща при її значній потужності (більше 50 м, як у Причорномор'ї), породи, прикриті четвертинними відкладами водно-льодовикового походження, тощо. Причому при генетико-еволюційному аналізі геосистем зовсім не обов'язково точно встановлювати місцеположення нижньої межі, достатньо лише вказати, в якій саме верстві гірських порід вона знаходиться. У цій верстві нижня межа має вигляд перехідної смуги, в межах якої геосистемні властивості природи поступово щезають. Взагалі, вплив верхніх геогоризонтів (аеро-, фіто- та педо-) на літосферу обмежується глибиною до кількох десятків метрів (зона гіпергенезу). Саме нижче цієї зони й доцільно визначати орієнтовне положення нижньої межі геосистем.
При аналізі міграційних потоків у геосистемі положення її нижньої межі визначається глибиною можливого проникнення мігруючої речовини. Ця глибина залежить від хімічних властивостей речовини-мігранта, характеру зони аерації (її фільтраційних особливостей, наявності ландшафтно-геохімічних бар'єрів тощо), глибини проникнення коренів рослин у грунт та інших факторів. У принципі при врахуванні усіх їх глибину міграції, а відтак і положення нижньої межі геосистеми можна визначити. Проте рівень сучасних знань про поведінку багатьох речовин у ландшафті недостатній, щоб бути цілком упевненим щодо точності та вірогідності таких оцінок. Виходячи з того, що у вертикальній міграції більшості речовин надзвичайно велику роль відіграють низхідні та висхідні потоки вологи, доцільно за нижню межу геосистеми прийняти рівень залягання ґрунтових вод. Лише впевнившись у тому, що мігруюча речовина не досягає цього рівня і акумулюється на певному ландшафтно-геохімічному бар'єрі, останній можна визнати за нижню межу геосистеми.
При балансових дослідженнях геосистеми за її нижню межу слід вважати рівень, нижче якого шари вже не охоплюються кругообігом певної речовини. Для водного балансу це здебільшого рівень ґрунтових вод. Для теплового балансу він збігається з рівнем, починаючи з якого щезає річна амплітуда температури ґрунту. Глибина його залежить від температуропровідності ґрунту та амплітуди температур на його поверхні. В Україні вона становить від 10 до 17—20 м. Для круговороту органічної речовини за нижню межу геосистеми виправдано прийняти межу між геогоризонтами які охоплюються процесами гуміфікації, та де такі процеси вже не відбуваються. Здебільшого вона знаходиться у верхньому шарі ґрунтоутворюючої породи або між нею та материнською породою, хоч в окремих геосистемах коріння рослин може сягати і глибших шарів, а деякі гризуни та дощові черви (не кажучи вже про мікроорганізми) здатні проживати на глибинах 6—8 м і більше.
Контрольні запитання:
1.Що визначає тип внутрішньогеосистемних відношень?
2.На які типи можна поділити всю множину внутрішньогеосистемних зв‘язків?
3.Які розрізняють загальні підходи структуризації геосистеми?
4.Що вважають геокомпонентами?
5.Що розуміють під геомасою?
6.Що є основою для виділення ґрунтових геогоризонтів?
7.Що є характерними особливостями верхніх меж геосистем?
8.Що слід вважати за нижню межу геосистеми?
ВЕРТИКАЛЬНІ СТРУКТУРИ ГЕОСИСТЕМИ:
МІЖЕЛЕМЕНТНІ ВІДНОШЕННЯ ТА ПРОЦЕСИ
(процесна ландшафтна екологія)
Основні питання:
1.Генетико-еволюційні відношення.
Антропічні аспекти генетико-еволюційних відношень.
2.Потік і трансформація енергії.
3.Потоки вологи.
Типологія потоку вологи.
4.Міграція та обмін мінеральних речовин.
5.Забруднення та самоочищення геосистем.
6.Продуційні процеси.
Концептуальні основи аналізу продуційного процесу в екосистемі.
6.1.Генетико-еволюційні відношення
Мета:вивчити генетико-еволюційні відношення, коротку історичну довідку, загальну схему та основні положення, антропічні аспекти, основні терміни.
План:
1.Коротка історична довідка.
2.Загальна схема та основні положення.
3.Антропічні аспекти генетико-еволюційних відношень
Коротка історична довідка.
Найбагатші традиції генетико-еволюційного аналізу природних систем були в російській, а згодом у радянській фізичній географії. В.В. Докучаєв та його учні (Г.Ф. Морозов, О.М. Краснов, І.М. Крашенінніков та ін.) надавали генетичним аспектам дослідження природи надзвичайної ваги, тому докучаївську географічну школу небезпідставно називають генетичною. Та якщо в ґрунтознавстві генетичний принцип дійсно послідовно дотримувався і розвивався, то в ландшафтознавстві багато в чому він був декларативним, на практиці спрощувався. Так, часто дослідження генетичної суті ландшафту зводилось до аналізу його геолого-геоморфологічної будови, яка буцімто генетично зумовлює всю його структуру. Такий підхід особливо яскраво виявився в працях М.А. Солнцева та прихильників його наукового напряму.
У класичному ландшафтознавстві генетичний підхід визнавався як провідний при виділенні ПТК усіх рангів, їх типології, при районуванні, вирішенні інших питань. Проте, незважаючи на гіпертрофовано велику увагу ландшафтознавців до генетико-еволюційних проблем, їх ґрунтовно розроблено не було. Це зумовило появу в 70-х роках критичних статей щодо генетичного принципу та його евристичного й практичного значення (Д.Л. Арманд, Ф.М. Мільков та ін.). Було наголошено, що генетичні зв'язки між геокомпонентами аж ніяк не вичерпують усієї багатогранності внутрішньо-геосистемних відношень, багато з яких практично незалежні від походження та особливостей розвитку геосистеми.
Загальна схема та основні положення.
Мета аналізу генетико-еволюційних відношень між складовими вертикальної структури геосистеми — визначити закономірності поєднання геокомпонентів один з одним та взаємозалежність їх окремих властивостей. Він зводиться до вирішення двох основних груп питань: 1) виявити закономірності формування даного поєднання геокомпонентів та характер зв'язків між ними; 2) оцінити ступінь генетичної залежності між геокомпонентами та їх елементами.
Загальна схема формування вертикальної структури геосистеми така. Утворення її починається з нуль-моменту—появи наземного твердого абіотичного субстрату. Цим моментом може бути вихід території з-під рівня моря, вивільнення її з-під льодовика, перекриття поверхні вулканічною лавою тощо. Геологічні породи, що вийшли на поверхню, ґрунтові води та приземний шар атмосфери вступають у взаємодію між собою, яка виражається у вивітрюванні гірських порід та зміні морфології рельєфу. Такі геосистеми мають примітивну вертикальну структуру, у якій немає біотичних компонентів. Проте вже на абіотичній стадії розвитку геосистеми до неї з атмосфери та з поверхневими водами надходять мікроорганізми, життєдіяльність яких спричиняє формування та поступове збільшення вмісту органічної речовини у верхньому шарі гірських порід. Внаслідок цього стає можливим ецезис (приживання на новому місці) лишайників, мохів, а згодом і вищих рослин. Їх спори та насіння заносяться до геосистеми практично з нуль-моменту, але на безорганічному субстраті проростати вони не могли. Популяційна структура первинних рослинних угруповань примітивна і цілком визначається фізико-хімічними особливостями субстрату, а також видовим складом рослинності прилеглих геосистем. Конкурентні та інші біотичні відношення між популяціями та їх окремими особинами виражені слабко.
З моменту виникнення фітоценозу геосистема вступає у біотичну стадію розвитку. Для неї характерні інтенсифікація гумусоутворення, заселення тваринами та формування зооценозу, більша швидкість сукцесійних змін ґрунту, рослинності та мікробоценозів, стабілізація рельєфоформуючих процесів, більша трансформація приземного шару атмосфери (зокрема, внаслідок все більшої транспірації). Якщо на абіотичній стадії розвитку зміни геокомпонентів та їх зв'язок загалом детерміновані та односпрямовані, то на біотичній відношення між геокомпонентами значно ускладнюються і набувають все більшої стохастичності (ймовірносності). Формується розгалужена мережа обернених, міжгеокомпонентних зв'язків, тобто переважно односпрямовані відношення змінюються двосторонніми. Так, розвиток ґрунту починає залежати не від властивостей гірських порід, атмосфери і ґрунтових вод, а й від популяційного складу та продуктивності рослинних угруповань. У свою чергу, зміни ґрунту зумовлюють і зміни рослинності, тваринного населення (особливо ґрунтової фауни), мікробоценозів, які знову-таки зумовлюють певні зміни ґрунтових процесів. За таких зв'язків визначення геокомпоненту, який є провідним у формуванні вертикальної структури геосистеми, лишається сенсу.
Важлива особливість еволюційних відношень на біотичній стадії формування геосистеми зумовлена зростаючою самостійністю розвитку окремих геокомпонентів у процесі їх еволюції. Так, на перших етапах сукцесії рослинних угруповань формування їх популяційної структури майже повністю визначається абіотичними факторами — кліматом та субстратом. Ці фактори ніби задають генеральний напрям можливих змін рослинності. У подальшому роль внутрішньогеокомпонентних процесів (між- та внутрішньовидової конкуренції, симбіозу та ін.) набуває все більшого значення, і популяційний склад біоценозів все більше визначається власне цими відношеннями. Вони до певної міри узалежнюють фітоценоз від ґрунту, клімату та геологічної будови. Аналогічно і грунт у процесі свого розвитку стає все менш залежним від материнської породи та клімату і в рамках деякого заданого ними діапазону змін може розвиватися відносно самостійно. З розвитком ґрунтового профілю геологічні породи також стають менш залежними від атмосфери, рослинності, мікроорганізмів. Це дає змогу в одних кліматичних умовах на одному типі і навіть виді ґрунту формуватися різним рослинним угрупованням, різним видам ґрунту на одній геологічній породі тощо.
Таким чином, відповідність геокомпонентів один одному жорстко не детермінована, її характер можна визначити як детерміновано-стохастичний. Він полягає в тому, що вплив певного геокомпонента на інший (наприклад ґрунту на рослинність) визначає деякий діапазон змін рослинності, за який вони не можуть вийти, тоді як у рамках цього діапазону формування рослинних угруповань може визначатися суто внутрішньоценточними або й чисто випадковими процесами (наприклад, на дерново-підзолистих піщаних ґрунтах не можуть формуватися букові ліси, типчаково-ковилові степи, а різні асоціації хвойних та й мішаних лісів тут можливі). Так само хоч і формування ґрунту обмежене характером рослинності, геологічною будовою, кліматом, рівнем та хімічним складом ґрунтових вод, ці обмеження не настільки значні, аби визначити єдино можливий вид ґрунту у даних умовах (на плоских лесових рівнинах при глибокому рівні залягання ґрунтових вод за однакових умов посушливого південно-степового клімату можуть формуватися чорноземи звичайні, різні за вмісту гумусу, потужністю, глибиною залягання карбонатів і навіть механічного складу, але тут неможливі сірі лісові, дерново-підзолисті та інші типи ґрунтів).
Генетико-еволюційні відношення між геокомпонентами визначають деякий спектр класифікаційних підрозділів одного геокомпонента, які можуть відповідати певному типу іншого геокомпонента (наприклад, набір видів ґрунту, які можуть відповідати певному типу гірських порід; набір рослинних угруповань, що можуть формуватися на даному виді ґрунту; видів ґрунтів — під даною рослинною асоціацією тощо). Цей набір може бути дуже широкий, проте з урахуванням впливу на двосторонній міжгеокомпонентний зв'язок певного третього геокомпонента (наприклад, впливу клімату на зв'язок ґрунтів і рослинності) суттєво звужується (фільтрується). Послідовне врахування впливу всіх геокомпонентів на двосторонній зв'язок якихось двох з них приводить до виділення лише тих можливих варіантів поєднання геокомпонентів, яке може скластися в умовах конкретної геосистеми.
Антропічні аспекти генетико-еволюційних відношень
Втручання людини в ландшафт призводить до розладу еволюційно зумовлених відношень між його геокомпонентами. Найбільш характерна в цьому плані заміна еволюційних зв'язків між ґрунтом та рослинністю у природних геосистемах на антропічно регульовані відношення між ними в агрогеосистемах. Зведення лісів та їх заміна на трав'яні агроценози призводить до формування протиприродних, «еволюційно-абсурдних» зв'язків між ґрунтом та рослинністю, тваринним населенням, мікробоценозами. Наприклад, з природно-еволюційної точки зору зовсім нехарактерні відношення між дерново-підзолистими ґрунтами та злаковою сільськогосподарською рослинністю (еволюційно ці ґрунти пов'язані з мішаними або хвойними лісами). Якщо грунт як більш інерційний геокомпонент може тривалий час зберігати свої еволюційно-генетично зумовлені риси при взаємодії з новим біоценозом, то самі ці біоценози самостійно існувати в геосистемі не можуть, еволюційно відштовхуються нею. Так, в Україні значні ареали сірих лісових ґрунтів постійно знаходяться під ріллею ще з XVI — XVII ст., проте свого генетичного типу вони не змінили. Натомість, існування агроценозів було можливим лише завдяки їх щорічному відновленню людиною. Припинення культивування агроценозів приводить до поступового налагодження еволюційних відношень у геосистемі, і її корінний фітоценоз відновлюється (у лісових геосистемах у середньому через 300—600, в трав'яних — через 40—80 років).
Господарська діяльність людини призводить не тільки до видозміни еволюційних відношень між ґрунтом та біокомпонентами, а й викликає більш глибоку трансформацію внутрішньогеосистемних генетично зумовлених зв'язків, торкаючись водного режиму, рельєфоформуючих процесів тощо. Так, у генетичному відношенні рельєф геосистем Причорноморської низовини акумулятивного походження, проте з масовим розоранням земель цього регіону, іригацією зараз тут переважають рельєфоутворюючі процеси, характерні не акумулятивним, а денудаційним рівнинам.
Надмірне намагання штучно законсервувати геосистему в її природному стані призводить до руйнування в ній генетико-еволюційних відношень. Так, у лучних та степових геосистемах еволюційно склалися відношення між рослинними угрупованнями та тваринами, за яких для нормального розвитку рослинності необхідне відчуження деякої частини її щорічної продукції травоїдними тваринами. Заповідання степових геосистем у режимі повного виключення стравлення та косіння призводить до досить швидкої деградації рослинних угруповань, аж до випадання едифікаторних видів. Тому щоб підтримати заповідні степи в близьких до природних станах, необхідно ввести режим стравлення, близький до природно-еволюційного.
Врахування генетико-еволюційних відношень у геосистемах має значення для конструювання стійких і ефективних ландшафтів. Так, на цьому принципі має базуватися оптимальний підбір порід дерев для інтродукції, обґрунтування екологічно безпечних сівозмін тощо.
6.2.Потік і трансформація енергії
Мета:вивчити потік і трансформацію енергії, коротку історичну довідку, загальну схему, антропічні аспекти, типологію, основні терміни.
План:
1.Коротка історична довідка.
2.Загальна схема.
3.Антропічні аспекти.
4.Типологія.
Коротка історична довідка.
Першою концептуальною моделлю потоків енергії в екосистемі була схема трофічних шляхів для прерії, запропонована В. Шелфордом у 1913 р. Через 10 років подібну модель для арктичної тундри розробили Ч. Елтон та В. Саммерхейз. Проте кількісних характеристик енергетичних потоків у цих схемах не було. По суті етапною слід вважати працю Р. Ліндемана (1942), який першим кількісно оцінив потік енергії по всій трофічній структурі екосистеми. Як модельну він обрав екосистему оз. Седар-Бог-Лейк (шт. Міннесота). Аналогічні дослідження, але більш точними методами здійснив у 1957 р. Г. Одум на оз. Сілвер-Спрінгс (шт. Флорида). Згодом подібні дослідження виконано і для наземних екосистем (Ю. Одум, 1960; А. Макфедьєн, 1963; Г.І.Дювіньйо, 1964 та ін.). В усіх цих схемах описувались далеко не всі енергетичні потоки в екосистемі, а переважно між її біотичними елементами. У географії ж перевагу віддавали дослідженню потоків енергії між абіотичними елементами; було встановлено основні закономірності формування структури радіаційного балансу різних типів ландшафтів (М.І. Будико, Ю.Р. Раунер та ін.). Синтетичний напрям аналізу енергетичних потоків, у якому охоплювались як біотичні, так і абіотичні елементи екосистеми, був пов'язаний з моделюванням продуційного процесу, в основі якого лежить потік і трансформація енергії. Першу таку модель запропонували японські вчені М. Монсі та І. Саекі в 1953 р., а протягом 70—80-х років розроблено багато інших, проте переважно для агроекосистем.
Значний емпіричний матеріал з енергетики геосистем отримано в результаті робіт по Міжнародній біологічній програмі в 70-ті роки. Це дало змогу скласти повні моделі потоків енергії для основних типів природних екосистем. З них найбільш досконалою та широко відомою стала модель екосистеми прерії США (експериментальної станції Поні), розроблена під керівництвом Г. Ван-Дайна та Дж. Інніса.
Загальна схема.
Основним джерелом енергії для багатьох процесів у геосистемі є сонячна. Порівняно з нею енергії до геосистеми від інших джерел надходить дуже мало (теплової енергії з надр Землі — 0,04% сумарної сонячної радіації, тектонічних рухів — 0,0005%). При цьому сонячна енергія і використовується в геосистемі найбільш ефективно: вона здатна трансформуватися в інші види енергії (теплову, хімічну, механічну), завдяки їй відбувається продукування біомаси, вологообіг, циркуляція повітряних мас тощо.
Загальну схему потоку і трансформації сонячної енергії в геосистемі наведено на мал. . На верхню межу атмосфери надходить 2 кал/см2 хв сонячної енергії. Проходячи крізь атмосферу, вона послаблюється атмосферними газами та пилом. При цьому ступінь послаблення залежить від довжини хвилі (частоти) світла. 3 екологічної точки зору, найбільш важливими обставинами диференційованого послаблення випромінювання є дві: 1) ультрафіолетове випромінювання (найбільш небезпечне для протоплазми) практично не проходить крізь озоновий шар, що й забезпечує можливість життя на планеті; 2) менше всього послаблюється видиме світло, що необхідне для фотосинтезу, а тому він може відбуватись і в похмурі дні.
Сумарна радіація (R на мал. ) складається з прямої (RІ) та розсіяної (RS). Співвідношення між ними залежить від географічного положення геосистеми, хмарності та інших кліматичних факторів. Це співвідношення визначає ряд похідних ландшафтно-екологічних факторів, зокрема — експозиційний. Чим більша частка прямої радіації в сумарній, тим більше значення має фактор експозиції схилу.
Частина сумарної радіації, досягаючи геосистеми, витрачається на ефективне випромінення в атмосферу RF та відбивається поверхнею геосистеми (відбита радіація RА). Величина останнього потоку визначається характером поверхні геосистеми (її альбедо) і суттєво змінюється в різних умовах: у дібровах — 0,14— 0,17, степах—0,20—0,23, на солончаках—0,35, засніженій поверхні —до 0,95.
Альбедо слід розглядати як інтегральний параметр вертикальної структури геосистеми, який визначає потік відбитої радіації з неї як з деякого нерозчленованого цілого. Насправді ж потік сонячної радіації, проходячи від верхньої межі геосистеми до поверхні ґрунту, значно змінює свою.-інтенсивність, спектральний склад та інші характеристики. Ці зміни визначаються геогоризонтною вертикальною структурою геосистеми, особливо потужністю та часткою геомас листя в окремих аерофітогоризонтах.
Ефективне випромінювання та відбита радіація втрачаються для геосистеми, а та частина сумарної радіації, що безпосередньо йде на різні процеси в геосистемі, називається її радіаційним балансом RВ. Більша його частина витрачається на випаровування (фізичне Е і транспірацію Т) та на турбулентну віддачу атмосфері Р, тобто на забезпечення вологообігу та прогрівання повітря геосистеми.

Мал. . Схема потоку енергії в геосистемі:
Сонячна радіація: R — сумарна, RІ — пряма, RS — розсіяна, RА—відбита; RЕ — ефективне випромінення; RВ — радіаційний баланс; Т — витрати тепла на транспірацію; Е — те ж на фізичне випаровування; Q — турбулентна віддача тепла атмосфері; Р — енергія на фотосинтез; Рb—енергія дихання рослин; Рa — чиста первинна продукція; Рm—втрата енергії з опадом; Рi — енергія у фітомасі; ВМ — енергія, накопичена в прирості біомаси; Z — енергія, що надходить до травоїдних тварин (консументів 1-го трофічного рівня); Zb —енергія дихання тварин; Zi—енергія синтезу нової зоомаси; Zт — енергія, що втрачається із загибеллю тварин; Zz — енергія, що переходить до наступного трофічного рівня; M — енергія мортмаси; Мb — енергія на дихання сапротрофів; Мc — енергія окислення мортмаси; Мm — енергія мінералізації мортмаси; Н — енергія, накопичена в гумусі
Витрати тепла на фотосинтез Р становлять дуже малу частку радіаційного балансу —в середньому 1,3 %. Проте її роль у геосистемі надзвичайно велика, бо власне вона зумовлює продуційний та інші важливі біотичні процеси. При фотосинтезі використовується фотосинтетичне активна радіація — ФАР, частка якої становить близько 45 % сумарної (40 % прямої та 62 % розсіяної). Рослинний покрив поглинає 90% світлової енергії ФАР, проте переважна його частина йде на транспірацію та регулювання температури рослин і лише 0,5—1,5 % —на фотосинтез. Ефективність фотосинтезу визначається гідротермічними умовами геосистем. Найбільша вона за максимальної теплозабезпеченості при оптимальному співвідношенні тепла й вологи. Такі природні умови характерні для екваторіальних лісів, де ефективність фотосинтезу досягає 4,5 % ФАР, або1,5 % сумарної радіації. У штучних умовах можна довести ефективність фотосинтезу до максимальної — 34 % ФАР.
Енергія Р, що пішла на забезпечення реакції фотосинтезу, витрачається на дихання рослин Рb (близько 50 %), а решта становить чисту первинну продукцію Рa — накопичення енергії в рослинній біомасі. З цієї енергії деяка її частина Рm з відмиранням рослин або їх окремих органів переходить до мортмаси, частина Pi залишається у фітомасі й зумовлює її щорічний приріст, частина Z разом з фітомасою, що поїдається тваринами, переходить до наступного трофічного рівня. Співвідношення між цими статтями енергетичних витрат залежить від багатьох факторів. З них особливо важливі: 1) видовий склад рослинного угруповання, 2) вікова структура популяцій, їх екологічний стан, 3) характер господарського використання, 4) чисельність та популяційний склад тваринного населення біоценозу.
Енергія Z, що надходить з фітомасою до травоїдних тварин, частково йде на потреби самих цих тварин, а частково переходить до тварин-хижаків. Ці тварини, в свою чергу, поїдаються хижаками вищих порядків, а відтак передають частину енергії і їм. Так формується трофічна структура геосистеми. В екології при її дослідженні прийнято виділяти трофічні рівні — сукупність організмів, які отримують енергію від рослин через однакове число організмів-посередників. Самі зелені рослини формують перший трофічний рівень — рівень продуцентів (або автотрофів); травоїдні тварини (зайці, олені тощо) — другий рівень (первинних консументів, або гетеротрофів 1-го порядку); хижаки — третій (рівень вторинних консументів, або гетеротрофів 3-го порядку) і так далі. Оскільки потік енергії від одного трофічного рівня до вищого супроводжується її втратами (до 95 %), кількість трофічних рівнів не може бути необмеженою і лише в енергетично багатих екосистемах може досягати 5—6 (екваторіальні ліси, морські планктонові екосистеми).
Насправді ж у реальних геосистемах трофічні відношення набагато складніші. Виділяються: 1) види, які живляться одночасно на скількох рівнях (наприклад, сови; лисиці, що поїдають не тільки дрібних ссавців, а й плоди рослин); 2) види, що час від часу змінюють свій рівень (наприклад, із зміною пір року, або в процесі старіння); 3) види, особини яких можуть поїдати одна одну (особливо це розвинуто у павуків та риб); 4) рослини, які поводяться як гетеротрофи, «поїдаючи» деяких тварин (венерина мухоловка, сарраценія, росичка). Тому більш реалістичне зображення потоку енергії між організмами на основі не трофічних рівнів, а трофічних ланцюгів (синонім — ланцюгів живлення) — графів, вершинами яких є популяції, а орієнтованими ребрами — потоки енергії, що передається з їжею від однієї популяції до, другої, яка цією популяцією може живитись.
Потік енергії Z від одного організму до іншого супроводжується її втратами. Їжа як рослинного, так і тваринного походження повністю організмом не засвоюється. Частина її виводиться з організму у вигляді екскрементів Ze, частина Zb витрачається на дихання тварин, а решта енергії Zi може йти на синтез нової біомаси (точніше—зоомаси) в результаті росту організму та розмноження. З цієї енергії частина Zт втрачається популяцією внаслідок загибелі її особин і далі включається в енергетичні потоки переробки мортмаси, а решта енергії Zz переходить до організмів-консументів вищих рівнів, де структура енергетичних потоків аналогічна. Величина потоку Z залежить насамперед від чисельності популяцій тварин та типу біому. Взагалі у трав'яних геосистемах відсоток наземної рослинної продукції, що споживається травоїдними тваринами, більший, ніж у лісових. У хвойних лісах він становить 2—3 %, широколистих— до 3, в степових геосистемах досягає 8 — 10, саванах Африки 28 — 60, а на культурних пасовищах США 30—45 %. Окремі популяції тварин у цьому відношенні відрізняються ще більше. Так, деякі насіннєїдні тварини можуть споживати до 100 % придатної для них їжі. Проте в середньому з одного трофічного рівня на наступний переходить не більше 5 — 15 % енергії. З цієї енергії травоїдні для утворення свого тіла та росту Zi використовують близько 1 — 10 % Zz, а решту Zь витрачають на дихання, з екскрементами Zе та відригуванням Zv причому частки цих втрат приблизно однакові.
Потоки та трансформація енергії, що міститься у відмерлих частинах рослин Рт, екскрементах консументів Zе та їх трупах Zт, ще не з'ясовані до кінця. Сукупність цих потоків прийнято називати детритним циклом геосистеми. Для багатьох типів геосистем роль цього циклу набагато вища, ніж трансформація енергії у їх гетеротрофному блоці. Це пояснюється тим, що частка енергії, яка включається в детритний цикл, становить 90 — 95 % первинної продукції порівняно з 5—10 %, що надходить від продуцентів до тварин-гетеротрофів.
Деяка частина енергії, що міститься у відмерлій фітомасі, втрачається через її окислення (переважно фотохімічне), в результаті чого виділяється СО2. У лучному степу Курського заповідника, наприклад, ця частка становить 4 % енергії відмерлої надземної фітомаси. У лісових геосистемах спостерігаються втрати енергії (10 — 30 %), зумовлені вимиванням дощовою водою органічних речовин із свіжовпалого листя. У схилових геосистемах значна частка енергії Рт втрачається з виносом її за межі геосистеми горизонтальними потоками води. Проте більшу роль у трансформації енергії в детритному циклі відіграють не абіотичні фактори, а сапрофаги (синоніми — редуценти, детритні консументи) — живі організми, що живляться неживим органічним матеріалом, який можна метаболізувати для отримання енергії. У наземних геосистемах сапрофаги споживають до 90 — 95 % усієї чистої первинної продукції. До них належать такі великі тварини, як грифи, ворони, краби, а також черви, кліщі, бактерії та гриби.
Потік енергії між організмами-сапрофагами теж має характер трофічного ланцюга (Ю. Одум увів для нього термін «детритний ланцюг»), проте трофічні відношення тут надто своєрідні і складніші, ніж серед гетеротрофних тварин. Деякі організми, як наприклад, дощові черви, безпосередньо споживають мертву органічну речовину, частина якої йде на їх розвиток, частина — на дихання, частина переробляється і виводиться з екскрементами, частина подрібнюється і стає більш придатною для їх подальшого розкладу грибами та бактеріями. Екскременти червів, а також інших сапрофагів знову споживаються — і представниками інших видів, і того самого. Черви, як і інші тварини-сапрофаги, разом з мертвою органічною речовиною споживають гриби і бактерії, що містяться в ній. Таким чином, трофічні відношення в детритному циклі являють собою складну мережу, в якій виділити трофічні ланцюги, досить складно, а трофічні рівні — практично неможливо.
Більша частина енергії в детритному циклі витрачається сапротрофами на дихання, а решта переходить спочатку в проміжні, продукти розкладу органічної речовини та неспецифічну органічну речовину ґрунту. У лучному степу в них залишається не більше 50 % енергії, що надходить з рослинними рештками і екскрементами. На наступному етапі трансформації цієї енергії до 70 % її втрачається на мінералізацію органічної речовини мікроорганізмами та до 25 % на їх дихання. Решта переходить у специфічні гумусові речовини — продукти гуміфікації. Таким чином, у гумус переходить не більше 5—10 % енергії рослинних решток.
Антропічні аспекти.
Практично будь-який аспект діяльності людини в геосистемі призводить до зміни у ній інтенсивності енергетичних потоків. Причому змінюються величина та співвідношення не тільки внутрішньогеосистемних потоків, а й вхідних та вихідних. Через забруднення атмосфери аерозолями дещо збільшується відбита радіація, тому до геосистем може надходити менше сумарної радіації. Так, смог здатний зменшити її на 30—40 %. Употоці сумарної радіації збільшується частка розсіяної, що призводить до деякого нівелювання експозиційних відмінностей
геосистем схилів.
У агрогеосистемах значне збільшення надходження енергії пов'язане із внесенням органічних добрив. Частина цієї додаткової енергії йде на формування врожаю, невелика частина консервується в гумусі, а значно більша (на схилах — до 60—70 %) непродуктивно втрачається геосистемою разом з виносом поверхневим та ґрунтовим стоком.
Трансформація характеру діяльної поверхні геосистеми внаслідок розорювання, зведення лісів, меліорації тощо призводить до зміни величини альбедо, а через неї — і до зміни структури радіаційного балансу. Так, альбедо геосистем із степовою рослинністю становить 19—23 %, а свіжозораних агроугідь на їх місці— 5 %; широколистих лісів 12—17 %, а полів зернових культур на їх місці — 22—28 %. Відповідно змінюється і частка відбитої радіації.
Зміни вертикальної структури геосистеми, пов'язані із зведенням природної рослинності, призводять до трансформації трофічної структури геосистем, а відтак — і потоків енергії між біотичними елементами. Найсуттєвішими тут є щорічні втрати енергії, накопичені геосистемою у фітомасі. Внаслідок цього зменшується; потік енергії, який надходить до детритного циклу — основи процесу продукування гумусу. Загалом трофічна структура агрогеосистем сильно спрощується, інтенсивність потоків енергії від продуцентів до первинних консументів значно зменшується, а сама сітка цих потоків стає менш розгалуженою. Це, зокрема, зумовлює низьку стійкість агрогеосистем порівняно з природними.
Типологія.
Ю. Одум (1986) запропонував такий поділ екосистем за основним джерелом надходження енергії: 1) природні, що отримують енергію тільки від Сонця; 2) природні, що отримують енергію від Сонця та додаткову енергетичну субсидію від інших природних джерел (заплави, схили, прибережні частини естуаріїв, маршів, конуси виносу тощо); 3) антропізовані, що отримують енергію від Сонця та додаткову субсидію від людини (найбільш типові— агроекосистеми); 4) промислово-міські системи, що отримують енергію палива (урбоекосистеми, індустріальні зони). У такому вигляді ця схема надто проста, проте за умови її розширення може бути корисною в багатьох відношеннях.
За кількістю енергії, яку отримує геосистема, розрізняються такі геосистеми (Мьойзель, 1965; В.Р. Волобуєв, 1973; Д.Г. Циганов, 1983): 1) мегатермні (радіаційний баланс RB>80 ккал/см2 на рік, поширені в екваторіальній зоні); 2) макротермні (RB 50—70, поширені в тропіках); 3) мезотермні (RВ 50—70, суб- і середземномор'я); 4) субмезотермні (RВ 40—50, неморальна зона); 5) субмікротермні (RВ 30-40, суббореальна зона); 6) мікротермні (RВ 20—30, бореальна зона); 7) нанотермні (RB<20 ккал/см2 на рік, поширені в суб- та аркт
| <== попередня сторінка | | | наступна сторінка ==> |
| Тема 4. | | | Потоки вологи |
|
Не знайшли потрібну інформацію? Скористайтесь пошуком google: |
© studopedia.com.ua При використанні або копіюванні матеріалів пряме посилання на сайт обов'язкове. |