РЕЗОЛЮЦІЯ: Громадського обговорення навчальної програми статевого виховання
ЧОМУ ФОНД ОЛЕНИ ПІНЧУК І МОЗ УКРАЇНИ ПРОПАГУЮТЬ "СЕКСУАЛЬНІ УРОКИ"
ЕКЗИСТЕНЦІЙНО-ПСИХОЛОГІЧНІ ОСНОВИ ПОРУШЕННЯ СТАТЕВОЇ ІДЕНТИЧНОСТІ ПІДЛІТКІВ
Батьківський, громадянський рух в Україні закликає МОН зупинити тотальну сексуалізацію дітей і підлітків
Відкрите звернення Міністру освіти й науки України - Гриневич Лілії Михайлівні
Представництво українського жіноцтва в ООН: низький рівень культури спілкування в соціальних мережах
Гендерна антидискримінаційна експертиза може зробити нас моральними рабами
ЛІВИЙ МАРКСИЗМ У НОВИХ ПІДРУЧНИКАХ ДЛЯ ШКОЛЯРІВ
ВІДКРИТА ЗАЯВА на підтримку позиції Ганни Турчинової та права кожної людини на свободу думки, світогляду та вираження поглядів
- Гідрологія і Гідрометрія
- Господарське право
- Економіка будівництва
- Економіка природокористування
- Економічна теорія
- Земельне право
- Історія України
- Кримінально виконавче право
- Медична радіологія
- Методи аналізу
- Міжнародне приватне право
- Міжнародний маркетинг
- Основи екології
- Предмет Політологія
- Соціальне страхування
- Технічні засоби організації дорожнього руху
- Товарознавство продовольчих товарів
Тлумачний словник
Авто
Автоматизація
Архітектура
Астрономія
Аудит
Біологія
Будівництво
Бухгалтерія
Винахідництво
Виробництво
Військова справа
Генетика
Географія
Геологія
Господарство
Держава
Дім
Екологія
Економетрика
Економіка
Електроніка
Журналістика та ЗМІ
Зв'язок
Іноземні мови
Інформатика
Історія
Комп'ютери
Креслення
Кулінарія
Культура
Лексикологія
Література
Логіка
Маркетинг
Математика
Машинобудування
Медицина
Менеджмент
Метали і Зварювання
Механіка
Мистецтво
Музика
Населення
Освіта
Охорона безпеки життя
Охорона Праці
Педагогіка
Політика
Право
Програмування
Промисловість
Психологія
Радіо
Регилия
Соціологія
Спорт
Стандартизація
Технології
Торгівля
Туризм
Фізика
Фізіологія
Філософія
Фінанси
Хімія
Юриспунденкция
Поділ ДНК зв'язуючих білків по їх структурі
Чинники транскрипцій і репресори.
На функціонування генів чинять вплив більшість білків. Причому цей вплив не укладається в ті відносно прості схеми оперонної регуляції, які реалізуються у бактерій.
Так, останніми роками виявлено і описано велика кількість білків, що мають властивості факторів транскрипцій або репресорів (або тих і інших відразу). При цьому вони вступають в різноманітні взаємини один з одним, а також з іншими речовинами, від чого залежить кінцевий вплив на активність генів.
Іншими словами, подобно складної сітки із сотен метаболічних перетворень, в клітинах еукаріот є не менш складна сітка регуляторних взаємовідношень. Повної картини цієї сітки поки немає, але окремі її фрагменти вже відомі.
В данную сітку входять не тільки білки, які безпосередньо взаимодіють з ДНК, але й велика кількість інших речовин — позаклітинні сигнальні речовини (в т. ч. гормони), клітинні і внутрішньоклітинні рецептори, внутрішньоклітинні посредники (медіатори). До того ж далеко не кожна регуляторна дія має своїм кінцевим ефектом зміну активності генів.
З усіх цих причин регуляторні взаємини у еукариот вимагають окремого розгляду, який ми розглянемо пізніше.
Тут же ми коротко відмітимо тільки ті моменти, які торкаються ДНК зв'язуючих білків, що виступають чинниками транскрипцій або репресорів. Таким чином, ви повинні мати на увазі, що йтиметься лише про кінцеву ланку складних регуляторних ланцюгів.
У різних білках ДНК-зв'язуючих білків часом зустрічаються схожі структурні елементи. За цією ознакою виділяють декілька родин таких білків. Проте потрібно відмітити, що не кожен ДНК-зв'язуючий білок вдається віднести до тої або іншої родини.
а) Білки, що містять мотив "спіраль-поворот-спіраль".

У цих білках (див.мал.) ДНК-впізнаваюча ділянка (складає невелику частину білку) включає дві α-спіралі, сполучених петлею. З них одна спіраль безпосередньо зв’язується з ДНК, специфічно взаємодіючи з визначеною послідовністю нуклеотидних пар.
У білці - дві однакові субодиниці, які одновременно беруть участь у взаємодії з ДНК. Це є можливим завдяки тому, що відповідна нуклеотидна послідовність ДНК є паліндромною, тобто, з урахуванням полярності ланцюгів, читається однаково з обох боків. (З паліндромами ми вже стикалися)
Особливо поширені подібні білки у прокаріот: маються на увазі білки-репресори.
Нижченаведені ж родини білків характерні, мабуть, лише для еукаріот.
б) Білки, що містять гомеодомени.
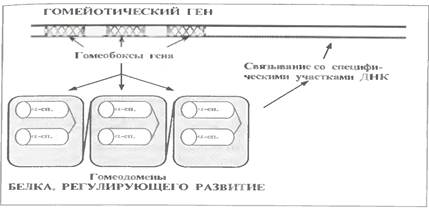
Ці білки продукти т. з. гомейотичних генів еукаріот, тобто генів, відповідальних за ембріональний розвиток.
За допомогою цих білків здійснюється керування розвитку шляхом включення одних і виключення інших генів.
Відмінна риса цих білків - наявність в них однотипних доменів, які називаються (по функції кодуючих генів) гомеодоменами (див.мал.).
Як відомо, домени - це фрагменти білкової субодиниці, що мають відносно самостійну третинну структуру. Іншими словами, в цьому випадку єдиний поліпептидний ланцюг при згортанні в третинну структуру утворює не одну велику, а дещо дрібніших глобулоподоб-них ділянок, пов'язаних між собою "лінкерними" ділянкими ланцюга.
Доменна організація зустрічається тільки у досить великих субодиниць (білків) - містять не менше 200 амінокислотних залишків. Фрагменти гена, що кодують різні домени білку, обов'язково розділені інтронами, тобто представляють собою різні екзони.
У разі гомейотических генів такі екзони називаються гомеобоксами.Отже, гомеобокс - це екзон гомейотичного гена, кодуючий гомеодомен відповідного білку.
Гомеодомен утворений приблизно 60 амінокислотними залишками. По структурі ж він нагадує белки-репресори прокаріот, тобто теж має мотив "спіраль-поворот-спіраль". І так само одна з α-спиралей цього мотиву специфічно взаємодіє з певними ділянками ДНК.
Гомеодомени різних білків цієї родини дуже схожі один з одним, що свідчить про спільність їх походження. Можливо, вони дійсно пішли від білків-репресорів прокаріот. Але при цьому кожен з гомеобілків робить дію не на окремий оперон, а на набагато більшу сукупність генів.
Дійсно, в процесі розвитку відбуваються масштабні зміни просторової структури великих фрагментів хромосом. Якісь з цих фрагментів переходять в стан гетерохроматину, інші - навпаки, в стан еухроматина.
Між іншим, звідси витікає, що і в гетерохроматині мають бути послідовності ДНК, доступні для впізнавання гомеодоменними білками.
в) Білки, що містять "цинкові пальці".

Білки цієї родини мають пальцеподібні структури (див.мал.). Кожна з останніх стабілізована атомом цинка, який утворює чотири координаційні зв'язки з двома залишками цистеїну і двома залишками гістидину пептидного ланцюга.
На зовнішній поверхні "пальця" знаходиться α-спіраль, що специфічно дізнається певну послідовність нуклеотидних пар ДНК.
Кількість "пальців" у різних білків неоднакова і сягає декількох десятків.
До цієї родини відносяться більшість регуляторних білок еукаріот. У тому числі - внутрішньоклітинні білки-рецептори стероїдних гормонів. Зв'язавши відповідний гормон, далі такий рецептор взаємнодіє "цинковими пальцями" з "своїм" енхансером у складі ДНК, що змінює транскрипційну активність певних генів.
г) Білки, що містять лейцинову"застібку".
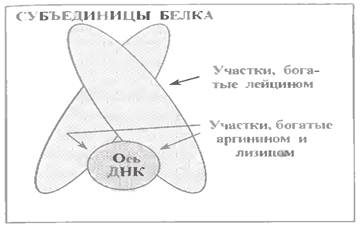
Ці білки (див.мал.) складаються з двох субодиниць, але останні можуть відрізнятися один від одного.
Головна особливість полягає в способі з'єднання субодиниць один з одним - це, в основному, гідрофобні зв'язки між залишками лейцину.
У ділянці ж зв'язування з ДНК білок багатий основними амінокислотами - аргініном і лізином. Це свідчить про те, що у взаємодії з ДНК важливе не стільки специфичне упізнавання білком певних послідовностей нуклеотидних пар (якщо таке взагалі має місце), скільки замикання іонних зв'язків з фосфатними групами ДНК.
За рахунок чого ж білок специфічно взаємодіє з потрібною ділянкою ДНК? Мабуть, усе визначається частково трьохмерною структурою цієї ділянки, а головне його білковим оточенням. Інакше кажучи, білок, що зв'язується, органічно вписується в ту просторову структуру (створювану макромолекулами), яка є в районі даного локуса ДНК. Іонні ж зв'язки просто закріплюють результат цього "вписування ".
До цієї родини відноситься більшість транскрипційнних чинників еукаріот. Тобто ті самі білки, які, як ми знаємо, взаємодіючи з ДНК, одночасно утворюють складні комплекси один з одним.
Як і гомеодоменні білки, дані білки, теж мають бактерійні "прототипи" - наприклад, Димерний білок САР (білковий активатор катаболізму).
Вищесказане підводить до наступного припущення. Чим тонша регуляція (тобто чим менше її масштаб або чим менше у регуляторного білку генів-мішеней), тим більше значення має просторова структура регульованої ділянки хромосоми.
| <== попередня сторінка | | | наступна сторінка ==> |
| Гени гемоглобіну | | | Загальні чинники транскрипції |
|
Не знайшли потрібну інформацію? Скористайтесь пошуком google: |
© studopedia.com.ua При використанні або копіюванні матеріалів пряме посилання на сайт обов'язкове. |