РЕЗОЛЮЦІЯ: Громадського обговорення навчальної програми статевого виховання
ЧОМУ ФОНД ОЛЕНИ ПІНЧУК І МОЗ УКРАЇНИ ПРОПАГУЮТЬ "СЕКСУАЛЬНІ УРОКИ"
ЕКЗИСТЕНЦІЙНО-ПСИХОЛОГІЧНІ ОСНОВИ ПОРУШЕННЯ СТАТЕВОЇ ІДЕНТИЧНОСТІ ПІДЛІТКІВ
Батьківський, громадянський рух в Україні закликає МОН зупинити тотальну сексуалізацію дітей і підлітків
Відкрите звернення Міністру освіти й науки України - Гриневич Лілії Михайлівні
Представництво українського жіноцтва в ООН: низький рівень культури спілкування в соціальних мережах
Гендерна антидискримінаційна експертиза може зробити нас моральними рабами
ЛІВИЙ МАРКСИЗМ У НОВИХ ПІДРУЧНИКАХ ДЛЯ ШКОЛЯРІВ
ВІДКРИТА ЗАЯВА на підтримку позиції Ганни Турчинової та права кожної людини на свободу думки, світогляду та вираження поглядів
- Гідрологія і Гідрометрія
- Господарське право
- Економіка будівництва
- Економіка природокористування
- Економічна теорія
- Земельне право
- Історія України
- Кримінально виконавче право
- Медична радіологія
- Методи аналізу
- Міжнародне приватне право
- Міжнародний маркетинг
- Основи екології
- Предмет Політологія
- Соціальне страхування
- Технічні засоби організації дорожнього руху
- Товарознавство продовольчих товарів
Тлумачний словник
Авто
Автоматизація
Архітектура
Астрономія
Аудит
Біологія
Будівництво
Бухгалтерія
Винахідництво
Виробництво
Військова справа
Генетика
Географія
Геологія
Господарство
Держава
Дім
Екологія
Економетрика
Економіка
Електроніка
Журналістика та ЗМІ
Зв'язок
Іноземні мови
Інформатика
Історія
Комп'ютери
Креслення
Кулінарія
Культура
Лексикологія
Література
Логіка
Маркетинг
Математика
Машинобудування
Медицина
Менеджмент
Метали і Зварювання
Механіка
Мистецтво
Музика
Населення
Освіта
Охорона безпеки життя
Охорона Праці
Педагогіка
Політика
Право
Програмування
Промисловість
Психологія
Радіо
Регилия
Соціологія
Спорт
Стандартизація
Технології
Торгівля
Туризм
Фізика
Фізіологія
Філософія
Фінанси
Хімія
Юриспунденкция
Глюкагон
Глюкагон - це одноланцюговий пептид (29 амінокислотних залишків), який утворюється в α-клітинах острівкового апарату підшлункової залози, а також в ШКТ (в кишечнику поряд з глюкагоном є низка глюкагонподібних сполук, які близькі за структурою і деякими властивостями до глюкагону).
Глюкагон синтезується у вигляді великого попередника - проглюкагону, який підлягає частковому протеолізу і перетворюється в активний гормон.
У плазмі глюкагон знаходиться у вільному стані, незв’язаному з транспортними білками. Саме тому він має короткий період напівжиття ≈ 5 хв. Інактивація відбувається в печінці під дією ферменту, який відщеплює від N-кінця пептиду дві амінокислоти.
Секреція глюкагону подавляється глюкозою. На секрецію гормону також впливають інші сполуки, такі як амінокислоти, жирні кислоти і кетонові тіла, гормони ШКТ і нейромедіатори.
Механізм дії цього гормону - мембранно-цитозольний, через активацію аденілатциклази і утворення цАМФ.
Біохімічні ефекти глюкагону протилежні ефектам інсуліну (це контрінсулярний гормон). Цей гормон викликає потужну мобілізацію джерел енергії за рахунок активації катаболізму вуглеводів і ліпідів.
Вплив на обмін вуглеводів спрямований на підвищення концентрації глюкози в крові. Основна “мішень” дії глюкагону - печінка, де він:
1) стимулює розщеплення глікогену (активує глікогенфосфо-рилазу) і інгібує глікогенез (гальмує активність глікоген-синтази); глюкагон не впливає на глікогеноліз у м’язах;
2) підвищує інтенсивність глюконеогенезу. Це досягається через активацію синтезу ключового ферменту фосфоенолпіруват-карбоксикінази (ФЕПКК) - цАМФ підвищує швидкість транскрипції гену ФЕПКК.
Вплив глюкагону на обмін ліпідів полягає в активації ліполізу. В адипоцитах гормон активує гормончутливу ліпазу, що сприяє підвищенню вільних жирних кислот в крові. Ці жирні кислоти можуть бути використані як джерела енергії або перетворюватися в печінці в кетонові тіла. В умовах інсулінової недостатності цей механізм є суттєвим у розвитку кетонемії.
6.4 Гормони наднирників
6.4.1 Гормони мозкового шару наднирників
У хромафінній тканині мозкового шару наднирників утворюються катехоламіни (адреналін - 85%, норадреналін).
Адреналін (епінефрин) - гормон, який секретується в наднирниках при стресі (концентрація його в крові зростає в 4-5 разів) і є медіатором в деяких синапсах. Встановлено також, що при стресі вміст в крові норадреналіну, який має переважно медіаторні властивості також зростає.
Катехоламіни синтезуються з амінокислоти тирозину. Перетворення тирозину в норадреналін і потім в адреналін має чотири послідовні етапи:
1 Гідроксилювання кільця (тирозин-гідроксилаза).
2 Декарбоксилювання (ДОФА-декарбоксилаза).
3 Гідроксилювання бічного ланцюга (дофамін-бета-гідрокси-лаза-ДБГ).
4 N-метилювання (фенілетаноламін-N-метилтрансфераза).
Послідовність реакцій синтезу виглядає таким чином:
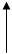

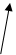

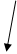



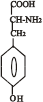
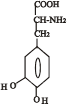

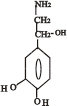
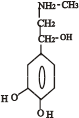
-ОН СО2 –ОН СН3
Тир ДОФА Дофамін Норадреналін Адреналін
(диоксифенілаланін)
Стадією, яка лімітує весь синтез є 1 - гідроксилювання Тир під дією тирозингідроксилази з утворенням ДОФА. У цьому ланцюгу реакцій також утворюється дофамін - катехоламін, який має переважно медіаторні властивості. Катехоламіни діють через два головні класи рецепторів: альфа-адренергічні і бета-адренергічні. Адреналін зв’язується з обома класами рецепторів. Норадреналін у фізіологічних концентраціях головним чином з альфа-рецептором.
Біохімічні ефекти адреналіну
Адреналін - це контрінсулярний гормон, дія якого спрямована насамперед на збільшення концентрації глюкози в крові. Крім того, цей гормон має катаболічну дію на обмін ліпідів.
Вплив адреналіну на обмін вуглеводів полягає в стимуляції глікогенолізу в печінці і м’язах. Це досягається шляхом актива-ції глікогенфосфорилази. У м’язах відбувається розщеплення глікогену до глюкозо-6-фосфату і далі до лактату. У м’язах розщеплення глікогену до глюкози не відбувається, тому що відсутня глюкозо-6-фосфатаза (яка розщеплює глюкозо-6-фосфат до глюкози). У печінці в результаті глікогенолізу утворюється вільна глюкоза, яка надходить у кров, що призводить до підвищення рівня глікемії. Це має велике значення для забезпечення інших тканин паливом в умовах стресу.
Вплив адреналіну на обмін ліпідів полягає в стимуляції ліполізу в жировій тканині через активацію ключового ферменту цього процесу - ТАГ-ліпази. У результаті в крові підвищується вміст вільних жирних кислот, що є додатковим джерелом метаболічної енергії.
Таким чином, сумарний біохімічний ефект адреналіну - це підготовка до ситуацій, в яких організм використовує значну кількість метаболічної енергії.
В інактивації катехоламінів беруть участь два ферменти:
1) моноамінооксидаза (МАО) - мітохондріальний фермент;
2) катехол-О-метилтрансфераза - фермент цитозоля.
Найбільша активність цих ферментів спостерігається в печінці і нирках.
Визначення концентрації катехоламінів в крові є важливим діагностичним підтвердженням діагнозу феохромоцитом. Феохромоцитоми - це пухлини мозкового шару наднирників, які секретують катехоламіни у високих концентраціях. У 90% випадків ці пухлини доброякісні. Як правило ці пухлини секретують адреналін і норадреналін, але переважно норадреналін. Дуже рідко переважаючим катехоламіном є дофамін (для злоякісних пухлин). Крім катехоламінів ці пухлини можуть продукувати серотонін, АКТГ, соматостатин, опіоїдні пептиди, кальцитонін та інші, що перешкоджає встановленню діагнозу.
Для феохромоцитом характерні гіперглікемія і глюкозурія, порушується толерантність до глюкози.
Механізм дії адреналіну
Адреналін - це гормон, для якого було вперше встановлено наявність мембранно-цитозольного механізму дії. При дії гормону на α2- і β-адренорецептори активується аденілатциклаза і синтезується цАМФ - вторинний посередник адреналіну, який запускає каскад метаболічних змін.
У разі зв’язування адреналіну з α1-адренорецептором активується фосфоінозитидний механізм, в якому роль месенджерів 


 виконують ІТФ, ДАГ і Са2+.
виконують ІТФ, ДАГ і Са2+.
Мозковий шар наднирників також продукує деякі нейропептиди - нейротензин, мен-енкефалін, β-ендорфін.
6.4.2 Гормони кори наднирників
Гормони, які синтезуються наднирниками з холестеролу поділяються на три групи: прегнанові (С21), андростанові (С19) і естранові (С18). До прегнанових стероїдів належать кортикосте-роїди і прогестини (які є проміжними метаболітами в синтезі багатьох стероїдних гормонів і це важливий стероїдний гормон яєчників і плаценти).
За біологічними властивостями кортикостероїди поділяють на:
- глюкокортикоїди;
- мінералокортикоїди.
Глюкокортикоїди
До глюкокортикоїдів належать кортизол, кортизон, кортикостерон, 11-дегідрокортикостерон. Найбільш активним є кортизол (гідрокортизон) і кортикостерон.
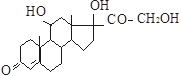
Кортизол
Після секреції в кров глюко- і мінералокортикоїди зв’язуються з білком транскортином (α-глобуліном, який синтезується в печінці). Мінералокортикоїди зв’язуються також з альбуміном. Лише 10% плазменних глюкокортикоїдів вільні і метаболічно активні. Активних вільних мінералокортикоїдів ≈ 30%.
У печінці відбувається інактивація глюкокортикоїдів. У результаті утворюються метаболіти з малою гормональною активністю і кон’юговані сполуки стероїдних гормонів з глюкуроновою і сірчаною кислотами. Більша частина їх екскретується нирками, менша з жовчю.
Глюкокортикоїди впливають на різні види метаболізму. Залежно від типу тканини вони можуть мати як анаболічну, так і катаболічну дію. Так, у печінці вони мають переважно анаболічний ефект, а в позапечінкових тканинах (ліпоїдній, м’язовій, сполучній) - це катаболічна дія.
Глюкокортикоїди беруть участь в регуляції обміну вуглево-дів, білків, впливають на проникливість клітинних мембран для глюкози і амінокислот, мають протизапальну і десенсибілізуючу дію.
Участь в обміні вуглеводів
Глюкокортикоїди - це гіперглікемічні гормони, тобто їх дія направлена на збільшення концентрації глюкози в крові. Це забезпечується за рахунок того, що гормони:
1) активують глюконеогенезу, що реалізується за рахунок стимуляції і координації декількох механізмів:
- індукують синтез ключових ферментів глюконеогенезу, у тому числі і ФЕПКК;
- індукують синтез ферментів, які в печінці каталізують реакції катаболізму амінокислот з подальшим їх перетворенням в глюкозу;
- стимулюють у позапечінкових тканинах катаболізм білків, який є джерелом амінокислот для глюконеогенезу.
2) гальмують поглинання і утилізацію глюкози в позапечінкових тканинах.
У печінці глюкокортикоїди діють як анаболічні гормони. Вони стимулюють не лише глюконеогенез. Глюкокортикоїди підсилюють глікогенез. Це відбувається шляхом активації ключового фермента - глікогенсинтази.
Участь в обміні ліпідів
Глюкокортикоїди підвищують мобілізацію ТАГ із жирового депо. У результаті в крові зростає концентрація гліцеролу і вільних кислот, які використовуються як джерела енергії. Крім того, гліцерол є додатковим субстратом для глюконеогенезу. В умовах інсулінової недостатності зростання концентрації ВЖК в крові є додатковим фактором, який стимулює кетогенез.
Надлишок глюкокортикоїдів стимулює ліполіз в одних частинах тіла (кінцівках) і ліпогенез - в інших (обличчя і тулуб)
Глюкокортикоїди здатні збільшувати ліполітичну дію сомато-тропіну і катехоламінів (пермісивний ефект глюкокортикоїдів). Крім того глюкокортикоїди мають незначну мінералокортикоїд-ну активність.
Участь в обміні білків
У печінці глюкокортикоїди мають анаболічний вплив на обмін білків і нуклеїнових кислот, в позапечінкових тканинах (м’язах, лімфоїдна, жирова, сполучна тканини) - катаболічний ефект. Молекулярний механізм анаболічної дії полягає в стимуляції транскрипції і відповідно трансляції специфічних білків (ферментів печінки, альбумінів плазми). Активація протеолізу білків у позапечінкових тканинах є джерелом амінокислот для синтезу глюкози в печінці і необхідна для активації саме глюконеогенезу.
Глюкокортикоїди беруть участь в функціонуванні захисних механізмів організму:
1) у високій концентрації (лікувальних дозах) гальмують імунологічну відповідь організму;
2) здатні гальмувати запальну реакцію. У фармакологічних дозах вони гальмують активність фосфоліпази А2. Цей фермент каталізує вивільнення арахідонової кислоти, яка є попередником синтезу простагландинів - гістогормонів - медіаторів запалення.
При аномальному збільшенні секреції глюкокортикоїдів виникає патологічний стан - хвороба (синдром) Іценко-Кушинга. Ця патологія розвивається при пухлинах надниркових залоз (синдром) або гіпофіза - підвищення продукції АКТГ (хвороба).
Основні прояви:
1) зниження толерантності тканини до глюкози;
2) гіперглікемія;
3) глюкозурія;
4) порушення ліпідного обміну - ожиріння з перевагою відкла-дання ліпідів у верхній частині тулуба, животі і обличчі;
5) затримка Na+ і важка гіпертензія.
Мінералокортикоїди
Це важливі гормони, які регулюють водно-сольовий обмін. Мінералокортикоїди, які синтезуються в наднирниках - це альдостерон, 11-дезоксикортикостерон (ДОК), 18-оксикортикостерон. Найбільш активним є альдостерон, в нормі лише він надходить у кров.
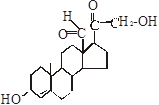
Альдостерон
Цей гормон регулює баланс в організмі Na+, К+, Cl- і води. Мішенями для альдостерону є клітини епітелію дистальних канальців нирок, в цитозолі і ядрі яких розташовані рецептори до гормону. При зв’язуванні альдостерону з рецепторами, комплекси, які утворюються, надходять в ядро і активують транскрипцію генів. Ці гени кодують інформацію про структуру транспортних білків плазматичних мембран епітелію канальців, які приймають участь в транспорті Na+.
Транспорт Na+ - це енергозатратний процес і потребує енергії АТФ. Альдостерон підвищує активність ферментів мітохондрій, які беруть участь в продукції АТФ. У результаті підсилюється реабсорбція Na+ і його протиіона з сечі в міжклітинну рідину і далі у кров.
Поряд з цим в обмін на Na+ підсилюється екскреція в сечу з епітелію канальців К+.
Таким чином, альдостерон забезпечує затримку Na+, Н2О (яка утримується Na+), Сl- в організмі і втрату з сечею К+.
Альдостерон також має протизапальну дію.
Регуляція секреції мінералокортикоїдів
Основним регулятором продукції альдостерону є ренін-ангіотензинова система. У процесі регуляції секреції беруть участь також К+, Na+, АКТГ.
Ренін-ангіотензинова система
Ренін-ангіотензин-альдостероновий механізм забезпечує регуляцію кров’яного тиску і електролітного обміну. Основним гормоном, який бере участь в регуляції альдостерону є ангіотензин ІІ. Схема його утворення наведена нижче.
Ангіотензиноген
↓ ренін
Ангіотензин І
↓ ангіотензин-перетворюючий фермент
Ангіотензин ІІ
↓ аміпептидаза
Ангіотензин ІІІ
↓ ангіотензинази
Продукти розпаду
За цією схемою ангіотензин ІІ утворюється з ангіотензину-гену - це α2-глобулін, який синтезується в печінці. Синтез ангіотензиногену підсилюють глюкокортикоїди і естрогени. Ангіотензиноген перетворюється в ангіотензин І під дією ферменту реніну, який синтезується в юкстагломерулярних клітинах нирок. Стимулами для виділення реніну є:
1) зниження кров’яного тиску;
2) втрата солей організмом;
3) зневоднення, втрата крові;
4) дія β-адренергічних агентів і простагландинів.
Під дією ангіотензин-перетворюючого ферменту (це гліко-протеїн, який знаходиться в легенях ендотеліальних тканинах і в плазмі крові) ангіотензин І перетворюється до ангіотензину ІІ. З ангіотензину ІІ утворюється ангіотензин ІІІ. У людини в плазмі крові концентрація ангіотензину ІІ в 4 рази вища, ніж ангіо-тензину ІІІ. Тому саме ангіотензин ІІ має основний вплив на секрецію альдостерону. У реалізації молекулярного механізму дії ангіотензину ІІ основна роль належить цАМФ.
Біохімічні і фізіологічні ефекти ангіотензину ІІ:
1 Активує синтез альдостерону і його секрецію (що призводить до затримки Na+, Cl- і Н2О).
2 Викликає звуження судин (що важливо в підтримці кров’яного тиску).
3 Викликає почуття спраги.
Гіперпродукція ангіотензину ІІ сприяє розвитку реноваску-лярної гіпертензії, серцевих набряків і порушення водно-сольо-вого обміну. Підвищення продукції мінералокортикоїдів спосте-ріггається при хворобі Конна (гіперальдостеронізм). При цьому захворюванні спостерігаються симптоми, які пов’язані з над-лишком альдостерону і гіпернатріємія, набряки, гіпертензія, під-вищена збудливість міокарда, знижений рівень реніну і ангіотензину ІІ.
Зниження секреції всіх кортикостероїдів (як глюкокортикої-дів, так і мінералокортикоїдів) спостерігається при гіперкорти-цизмі - хвороба Аддісона (бронзова хвороба). При цьому захворюванні пошкоджено більше 90% клітин кори надирників, які продукують кортикостероїди. Друга назва цієї патології - бронзова хвороба - пов’язана з тим, що поступово розвивається гіперпігментація - шкіра стає рівномірно бронзового кольору. Вважають, що це зумовлено надмірною секрецією МСГ. Включається компенсаторний механізм, який полягає в підвищенні секреції АКТГ і відповідних продуктів процесингу ПОМК (в тому числі і МСГ).
До біохімічних і фізіологічних змін при хворобі Аддісона належать: гіпоглікемія, надмірна чутливість до інсуліну, невитривалість до стресу, втрата ваги, анорексія, слабкість, низький кров’яний тиск, зменшення швидкості клубочкової фільтрації, гіпонатріємія, гіперкаліємія.
| <== попередня сторінка | | | наступна сторінка ==> |
| Гормони периферичних залоз | | | Гормони статевих залоз |
|
Не знайшли потрібну інформацію? Скористайтесь пошуком google: |
© studopedia.com.ua При використанні або копіюванні матеріалів пряме посилання на сайт обов'язкове. |