РЕЗОЛЮЦІЯ: Громадського обговорення навчальної програми статевого виховання
ЧОМУ ФОНД ОЛЕНИ ПІНЧУК І МОЗ УКРАЇНИ ПРОПАГУЮТЬ "СЕКСУАЛЬНІ УРОКИ"
ЕКЗИСТЕНЦІЙНО-ПСИХОЛОГІЧНІ ОСНОВИ ПОРУШЕННЯ СТАТЕВОЇ ІДЕНТИЧНОСТІ ПІДЛІТКІВ
Батьківський, громадянський рух в Україні закликає МОН зупинити тотальну сексуалізацію дітей і підлітків
Відкрите звернення Міністру освіти й науки України - Гриневич Лілії Михайлівні
Представництво українського жіноцтва в ООН: низький рівень культури спілкування в соціальних мережах
Гендерна антидискримінаційна експертиза може зробити нас моральними рабами
ЛІВИЙ МАРКСИЗМ У НОВИХ ПІДРУЧНИКАХ ДЛЯ ШКОЛЯРІВ
ВІДКРИТА ЗАЯВА на підтримку позиції Ганни Турчинової та права кожної людини на свободу думки, світогляду та вираження поглядів
- Гідрологія і Гідрометрія
- Господарське право
- Економіка будівництва
- Економіка природокористування
- Економічна теорія
- Земельне право
- Історія України
- Кримінально виконавче право
- Медична радіологія
- Методи аналізу
- Міжнародне приватне право
- Міжнародний маркетинг
- Основи екології
- Предмет Політологія
- Соціальне страхування
- Технічні засоби організації дорожнього руху
- Товарознавство продовольчих товарів
Тлумачний словник
Авто
Автоматизація
Архітектура
Астрономія
Аудит
Біологія
Будівництво
Бухгалтерія
Винахідництво
Виробництво
Військова справа
Генетика
Географія
Геологія
Господарство
Держава
Дім
Екологія
Економетрика
Економіка
Електроніка
Журналістика та ЗМІ
Зв'язок
Іноземні мови
Інформатика
Історія
Комп'ютери
Креслення
Кулінарія
Культура
Лексикологія
Література
Логіка
Маркетинг
Математика
Машинобудування
Медицина
Менеджмент
Метали і Зварювання
Механіка
Мистецтво
Музика
Населення
Освіта
Охорона безпеки життя
Охорона Праці
Педагогіка
Політика
Право
Програмування
Промисловість
Психологія
Радіо
Регилия
Соціологія
Спорт
Стандартизація
Технології
Торгівля
Туризм
Фізика
Фізіологія
Філософія
Фінанси
Хімія
Юриспунденкция
Кістяк голови включає мозковий і лицьовий череп.
Мозковий череп утворять кістки: парні – тім’яні і височні та непарні-потилична, клиноподібна, лобна, ґратчаста.
Клиноподібна кістка розташована в підставі черепа. У ній розрізняють тіло, на верхній поверхні якого знаходиться турецьке сідло (Рис.31,Б). У його поглибленні розташовується гіпофіз – залоза внутрішньої секреції. З боків від тіла клиноподібної кістки знаходяться малі і великі крила.
У ґратчастій кістці розрізняють лабіринти, що утворять середню,верхню носові раковини, і пластинки: очноямкову, що беруть участь в утворенні очниці, перпендикулярну – формуючу перегородку носа, ґратчасту, через яку проходять волокна нюхового нерва (Рис.31,А).
Лицьовий череп утворений верхньощелепними,носовими, слізними, вилицевими, піднебінними, нижніми носовими раковинами, лемешем,нижньою щелепою і під’язичної кістками. У черепі є тільки одна рухливо – з’єднана кістка – нижня щелепа.
У лобній, скроневій, клиноподібній і верхньощелепній кістках є пазухи, що відкриваються в носову порожнину й у такий спосіб сполучаються з зовнішнім середовищем. Завдяки наявності цього з’єднання пазухи пневматизовані, тобто заповнені повітрям.
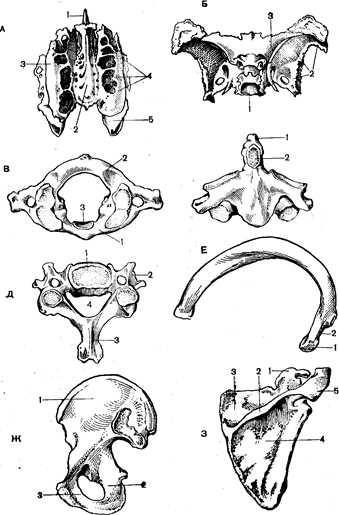
Рис. 31. Кістки кістяка людини:
А – ґратчаста (вид зверху): 1 – перпендикулярна пластинка;
2 – продірявлена; 3 – очноямкова; 4 – лабіринти; Б – клиноподібна:
1 – тіло; 2 – велике крило; 3 – мале крило; В – атлант: передня (1) і задня (2) дуги; 3 – суглобна поверхня для зубовидного відростка; Г – епістрофей: 1 – зубовидный відросток; 2 – суглобна поверхня; Д – шийний хребець: 1 – тіло; 2 -поперечний і 3 – остистий відростки; 4 – хребетний отвір; Е – ребро: 1 – голівка; 2 – шийка; Ж -тазова кіста: 1 – здухвинна; 2 – сіднична; 3 – лонна; З – лопатка: 1- дзьобуватий відросток; 2 – остюк;3 – надвістюкова яма;
4 – підвістюкова яма; 5 – акроміон.
Очниця утворюється кістками: зверху – лобною і клиноподібною, знизу – верхньощелепною і вилицевою, зовні – клиноподібною, лобною і вилицевою, із внутрішньої сторони – слізною і ґратчастою.
У порожнині носа містяться три носові раковини: верхня, середня і нижня, між ними розташовуються носові ходи, куди відкриваються пазухи.
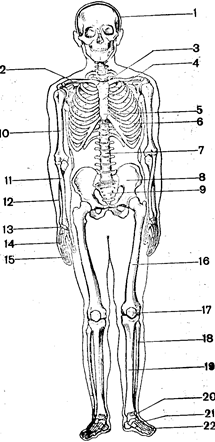
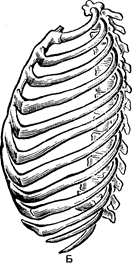
Кістяк тулуба складається з хребта і грудної клітини (Рис.32 ). Хребет включає 33–34 хребці:
|
Рис. 32. Кістяк людини:
1 – череп, 2 – ключиця, 3 – рукоятка;
4 – тіло і мечоподібний (5)відросток
грудної кістки: 6 – грудна клітина; 7 – хребет; 8 -тазові кістки; 9 – крижова кістка; 10 – плечова кістка; 11- променева кістка; 12 – ліктьова кістка; 13 – зап’ястя; 14 – п’ястя; 15 – фаланги пальців; 16 – стегнова кістка; 17 – надколінок; 18 – мала гомілкова кістка; 19 – гомілка;
20 – предплюсна; 21 – плюсна;
22 – фаланги пальців.
7 шийних, 12 грудних, 5 поперекових, 5 зрослих крижових і 4–5 зрослих куприкових. Відповідно виділяють шийний, грудний, поперековий, крижовий і куприковий відділи хребта.
Кожен хребець складається з тіла, дуги і відростків(Рис.31, Д). Розрізняють непарний остистий відросток, парні поперечні, верхні і нижні суглобні відростки, що відходять від дуги. Між задньою поверхнею тіла хребця і його дугою знаходиться хребетний отвір. Хребетні отвори всіх хребців утворюють хребетний канал, у якому розташований спинний мозок. На верхній і нижній границі дуги і тіла хребця знаходяться вирізки. Вони між верхнім та нижнім хребцями формують міжхребцеві отвори, у яких містяться спінальні ганглії.
Хребці різних відділів хребта мають деякі відмінності. Так, перший шийний хребець, називаний атлантом, має форму трохи витягнутого кільця (Рис.31,В). На його передній поверхні знаходиться суглобна ямка для зчленування з другим хребцем.
Другий хребець епістрофей – має зубовидный відросток, за допомогою якого здійснюється зчленування з першим хребцем (Рис.31,Г). Навколо зубовидного відростка здійснюється обертання атланта разом з черепом.
Грудні хребці мають зверху і знизу бічної поверхні тіла реберні напівямки для зчленування їх з голівкою ребра. Шийні і поперекові хребці мають поперечнореберні відростки.
Грудна клітина утвориться грудиною і 12 парами ребер. Ребро – вигнута пластинка – має голівку, шийкуі горбок. Голівкою і горбком ребра зчленовуються з грудними хребцями. Передні кінці ребер хрящові. Ребра з I по VII зчленовуються з грудиною, VIII–IX приєднуються до ребра, що знаходиться вище, кінці X, XI і XII пар ребер знаходяться в м’язах живота.
Грудина складається з рукоятки, тіла і мечоподібного відростка. Рукоятка з’єднується з ключицею першою парою ребер, II–VII пари ребер зчленовуються з тілом грудини.
Кістяк кінцівки складається з кістяка вільної кінцівки і кістяка пояса.
Пояс верхніх кінцівок містить у собі парні кістки лопатки і ключиці. Лопатка має ость, що закінчується плечевим відростком – акроміоном. Він утворює суглоб із ключицею. На одному з кутів лопатки міститься суглобна западина для зчленування з голівкою плечевої кістки.
Кістяк вільної верхньої кінцівки складається з, кісток передпліччя і кисті. Плечева кістка зчленовується своєю голівкою з лопаткою, а в нижній своїй частині утворить ліктьовий суглоб з кістками передпліччя: ліктьовий, розташованої по лінії мізинця, і променевий по лінії великого пальця. Нижній кінець променевої кістки утворить променезап’ястковийсуглоб із трьома кістками верхнього ряду зап’ястя. Кисть утворять кістки зап’ястя, п’ястя і фаланг пальців. Зап’ястя складається з 8 кісток, розташованих у два ряди. Верхній ряд включає човнувату, напівмісячну, тригранну і горохоподібну кістки, а нижній – кістку-трапецію,трапецієподібну, голівчату і гачкувату.
П’ясток утворить п’ять трубчастих кісток. Кістяк пальців складається з фаланг: другий – п’ятий пальці мають по три фаланги, а перший палець – дві.
Пояс нижніх кінцівок утворюють парні тазові кістки і криж. Кожна тазова кістка складається із здухвинної, сідничної і лонної. У місці їх сходження утвориться кульшівка, куди входить голівка стегна, формуючи кульшовий суглоб. На поверхні здухвинної кістки міститься суглобна поверхня для зчленування з крижем. Права і ліва лонні кістки з’єднуються попереду, утворюючи напівсуглоб.
Кістяк вільної нижньої кінцівки складається зі стегна, гомілки і ступні. Стегнова кіста в нижньому епіфізі має латеральний і медіальний виростки. Виростки забезпечені суглобними поверхнями, з якими зчленовуються надколінокі гомілка, утворюючи колінний суглоб.
Кістяк голінки складається з гомілки та малої гомілкової кісток. У верхньому кінці гомілки містяться два виростки, що зчленовуються з виростками стегна. Нижче і зовні бічного виростка гомілки розташована суглобна площина для зчленування з малою гомілковою кісткою. Нижній кінець гомілки з’єднується з таранною кісткою, що на зовнішній своїй стороні має суглобну поверхню для з’єднання з поверхнею малою гомілковою кісткою. Нижні кінці малої гомілкової кістки і гомілки зростаються нерухомо, між ними і таранною кістою формується скоковий суглоб.
Ступня складається з передплесно, плесно і фаланг пальців. Кістки передплесно розташовані в два ряди. Верхній ряд включає таранну і п’яткову кістки, нижній – першу, другу, третю клиноподібні і кубоподібну. Між цими двома рядами розташована човникова кістка. Таким чином, усього передплесно включає сім кісток. Плесно містить п’ять трубчастих кісток. Кістяк першого пальця складається з двох фаланг, а другого-п’ятого – із трьох.
БУДОВА І ВЛАСТИВОСТІ КІСТЯКОВИХ М’ЯЗІВ
Кістякові м’язи активно беруть участь в організації руху. Будь-яка рухова реакція організму здійснюється при участі м’язів, що, перетворюючи кістяк у систему важелів, сприяють переміщенню тіла в просторі.
В усіх м’язах розрізняють сухожильну голівку –початокм’язу, черевце, що складається з м’язових волокон, і кінець м’язу, названий сухожиллям. Звичайно м’яз прикріплюється до двох чи декількох кісток, які утворюють суглоб, що дозволяє їй при скороченні робити той чи інший рух у даному суглобі. Існують м’язи, у яких між прикріпленням її початку і кінця розміщені декілька суглобів. При такому характері прикріплення м’язу її скорочення викликає одночасний рух у всіх цих суглобів.
М’язи можуть бути простими і складними. У складних м’язах на відміну від простих черевце утворюється декількома голівками, котрі, починаючись від різних кісткових точок, потім зливаються разом (двоголова, триголова і чотириголова). Подібно цьому, сухожилля м’язу може поділятися на декілька частин і прикріплюватися до різних кісток. Місцем прикріплення м’язів, крім кісток, може бути шкіра, очне яблуко та ін.
Поверхня м’язу покрита фасцією, утвореною щільною сполучною тканиною. У місцях з’єднання двох сухожиль, чи сухожилля і кістки утворюються сполучнотканинні синовіальні сумки, у яких міститься невелика кількість рідини, що зменшує тертя рухливих поверхонь.

А Б В Г
Рис. 33. Розташування волокон у різних м’язах:
А – веретеноподібний; Б – одноперистий; В – двоперистий; Г – широкий.
У місцях проходження сухожиль у кістковому каналі вони покриваються синовіальнимипіхвами, усередині яких також міститься невелика кількість рідини, що, знімаючи тертя, полегшує рух.
М’язи класифікують по їх формі і функції. У залежності від форми м’язи поділяють на широкі (м’язи тулуба і поясів кінцівок), довгі (м’язи кінцівок), короткі(між хребцями), кругові (навколо отворів тіла). По функції розрізняють м’язи – згиначірозгиначі, що приводять і відводять м’язи, а також м’язи, що обертають всередину та назовні.
У довгих м’язах волокна можуть мати: 1) рівнобіжне розташування, орієнтоване уздовж довгої осі м’язів; 2) рівнобіжне розташування по відношенню один до одного і косе щодо довгої осі черевця; 3) косе розташування по відношенню один до одного і до довгої осі черевця. Відповідно розташуванню волокон розрізняють м’язи веретеноподібні, напівперисті і перисті. Напівперисті і перисті м’язи мають волокна більш короткі, ніж веретеноподібні, тому розмах рухів при їхньому скороченні менше. У широких м’язах волокна можуть розташовуватися паралельно (ромбовидні м’язи), радіально і віялоподібно (великий грудний м’яз). М’язи, у яких волокна розташовані радіально, можуть скорочуватися й у цілому, і окремими своїми частинами в напрямках, що перетинають різні осі руху в суглобі. Тому вони знаходяться в основному в ділянки кулястих суглобів, що відрізняються великою рухливістю.
У залежності від місця розташування м’язів їх поділяють на м’язи голови, шиї, тулуба (грудей, живота, спини), м’язи верхніх, нижніхкінцівок.
М’язи голови поділяють на жувальні і мімічні. Жувальні м’язи забезпечують рухи нижньої щелепи, беруть участь в акті жування; мімічні м’язи, прикріплюючись до шкіри обличчя, зміщають її при своєму скороченні, що і лежить в основі мімічних рухів: зморщування брів, піднімання й опускання кутів рота і т.д.
М’язи шиї здійснюють її згинання і рух голови, опускають нижню щелепу, піднімають ребра, беруть участь у диханні, зміщують під’язичну кістку і гортань, можуть фіксувати під’язичну кістку і тим сприяти виникненню звуків у гортані.
М’язи грудей, що розташовані поверхнево, рухають плечовий пояс і рукуі; що знаходяться більш глибоко, скорочуючись, здійснюють дихальний акт.
М’язи живота сприяють видиху, викликають згинання хребта вперед, убік і поворот його навколо подовжньої осі. Вони утворюють стінку черевної порожнини – черевний прес, сприяють виведенню сечі, калу і т.д.
М’язи спини, розташовані поверхнево, викликають рух руки, поясу верхніх кінцівок, розгинання голови, фіксування лопатки.
Більш глибоко розташовані м’язи спини беруть участь у дихальних рухах, викликають розгинання хребта, нахил його убік і обертання, розгинання й обертання голови, забезпечують фіксування хребта.
Структура м’язовихволокон. Структурною одиницею кістякових м’язів є поперечносмугасте м’язове волокно – скорочувальне багатоядерне утворення. Діаметр волокна коливається від 12 до 70 мкм, а довжина може досягати декількох сантиметрів. Зовні поперечносмугасте м’язове волокно покрите оболонкою – сарколемою. Усередині волокна знаходяться всі звичайні для клітини компоненти: цитоплазма, що у м’язовому волокні називається саркоплазмою, мітохондрії, саркоплазматичний ретикулум і т.д. У поперечносмугастому м’язовому волокні може бути більш ста ядер. Специфічні структури представлені в м’язовому волокні міофибрилами – тонкими нитками, що тягнуться від одного кінця поперечносмугастого м’язового волокна до іншого. Діаметр міофибрили складає 0,5–2 мкм. Кожна міофибрила складається з дуже тонких волоконець – протофібрил – різної довжини і товщини. Протофібрили мають характерне упорядковане розташування. Уздовж усієї міофібрили на відстані 2–3 мкм один від одного розташовані смужки, що позначаються буквою z.
Ділянку міофібрили між двома сусідніми смужками називають саркомером. Отже, уся міофібрила складається з повторюваних саркомерів. Усередині кожного саркомера тонкі протофібрили не доходять один до одного, тобто кінці їх не з’єднуються, залишаючи простір Н. Між двома поруч розташованими тонкими протофібрилами знаходяться товстіпротофібрили, що займають середнє положення в саркомері. Вони скріплені сіткоподібною структурою, представленою темною смугою М в центрі диска Н. Ділянку міофібрили, у якій розташовані товсті протофібрили, позначають диском Л. З двох сторін від нього розташовуються диски І, що складаються з тонких протофібрил. Диск А виглядає при розгляданні в мікроскоп більш темним, він неоднаково переломлює світло в подовжньому і поперечному напрямках, унаслідок чого його називають анізотропним (звідси і позначення його буквою А). Диск І ізотропний, він володіє не настільки вираженою анізотропією і виглядає більш світлим. Чергування в міофібрилах темних і світлих дисків додає м’язовому волокну поперечну смугастість, у зв’язку з чим м’язи, що складаються з таких волокон, називають поперековопосмугованими.
Товсті протофібрили складаються з білку міозину, а тонкі – актину і тропоміозину. Актин складає 20% від загальної кількості структурних білків м’язів, він позбавлений ферментативних властивостей. На долю міозину приходиться 60% білку м’язів. Міозин має здатність розщеплювати АТФ.
Протофібрили розташовані гексагонально: міозинові нитки розташовані по кутах шестикутника, у центрі якого також проходить міозинова нитка. Актинові нитки знаходяться в трикутнику між міозиновими. Товсті волокна забезпечені містками, що відходять від товстих протофібрил у різних площинах через кожні 60–70 Å.
Поперековопосмуговане м’язове волокно має складну структуру внутрішніх мембран – саркоплазматичний ретикулюм. Саркоплазматичний ретикулюм представлений системою канальців, пухирців, цистерн, розташованих у визначеному порядку й оточених мембранами. Порожнини ретикулюма розташовані між міофібрилами, утворити не сполучені між собою відсіки (Рис. 35). Вважають, що саркоплазматичний ретикулюм є транспортною системою м’язового волокна і бере участь у проведенні збудження від поверхневої мембрани до міофібрилам.
Механізм м’язового скорочення і розслаблення. В основі м’язового скорочення лежить переміщення ниток актину відносно ниток міозину.
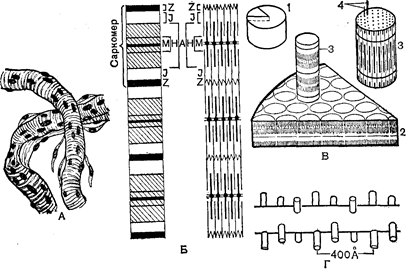
Рис. 34. Макро- і мікроструктурапоперековопосмугованого м’язу:
А – м’язове волокно; Б – схема будови міофібрили; В – схема будови ділянки поперековопосмугованого м’язу: 1- м’язове волокно; 2 – ділянка м’язового волокна; 3 – міофібрила; 4 – протофібрили; Г – поперечні містки між
тонкими і товстими протофібрилами.
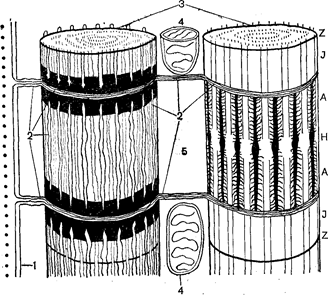
Рис.35. Схема структури саркоплазматичного ретикулюма:
1 – сарколема; 2 – саркоплазматичний ретикулюм;
3 – міофібрили; 4 – мітохондрії; 5 – саркоплазма.
Рис. 36. Зміни взаєморозташування тонких і товстих
протофібрил при скороченні волокна: 1- міозинові нитки; 2 – актинові нитки.
|
Нитки актину рухаються, як по тунелю, між міозиновими фібрилами, за рахунок чого коротшає ширина смужки Н(Рис. 36). Таке вклинювання актинових ниток між міозиновими приводить до зменшення довжини м’язу. Скорочення, що супроводжується зміною довжини, називають ізотонічним. При наявності якоїсь перешкоди скороченню, наприклад, коли до м’язу підвішений великий вантаж, він напружується, але довжина його не міняється. Такий тип скорочення, що здійснюється при незмінній довжині, називають ізометричним. Виявилося, що в основі цього виду скорочення лежить той же самий механізм, але в цьому випадку одні саркомери скорочуються, інші розтягуються, що забезпечує відсутність зміни довжини волокна.
У процесі ковзання ниток містки утворять зв’язку зі специфічними центрами молекул актину. Кожен місток може зчепитися з одною актиновою ділянкою, просунути нитку актину на якусь відстань, потім від’єднатися від нього і ввійти в контакт із наступним містком. Енергію для переміщення ниток дає АТФ при її розщепленні.
Ферментом, що розщеплює АТФ у м’язах, є міозин, активність якого різко зростає при з’єднанні з актином і утворенні білку актоміозину. Утворення актоміозину здійснюється тільки в присутності Са++. При введенні Са++ усередину м’язового волокна виникає його скорочення. Гранична концентрація Са++ складає 5 х 10 -7 моль, що значно менше, ніж у м’язовому волокні. У стані спокою скорочення м’язу не виникає. Це пояснюється тим, що Са++ знаходиться усередині саркоплазматичного ретикулюма. Для звільнення Са++ необхідна деполяризація мембран ретикулюма.
|
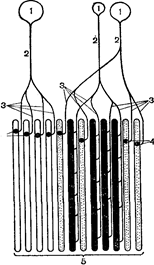
Рис. 37. Схематичне зображення рухової одиниці:1 – мотонейрони; 2 – аксони мотонейронів; 3 – розгалуження аксонів; 4– нервово – м’язові синапси; 5 – м’язові волокна.
Весь ланцюг подій, що відбуваються при м’язовому скороченні, можна представити в такий спосіб: у нервово – м’язовому синапсі під впливом імпульсів, що надходять по нервовому волокну, відбувається виділення ацетилхоліну, що деполяризує мембрану м’язового волокна. Виникаючий унаслідок цього імпульс поширюється по мембранах саркоплазматичного ретикулюма. Зміна потенціалу мембрани ретикулюма супроводжується збільшенням її проникності для Са++ і виходом його в міжфібрилярний простір. Са++ сприяє утворенню актоміозину, під впливом якого розщеплюється АТФ. Енергія, що звільняється при цьому, використовується для ковзання ниток. Ця теорія одержала назву теорії ковзних ниток (Хакслі А.).
Розслаблення м’язу пов’язане зі зворотним надходженням Са++ у саркоплазматичний ретикулюм, що відбувається при участі активних механізмів, пов’язаних з роботою іонних насосів.
Функціональні властивості кістякових м’язів. У природних умовах діяльності організму скорочення м’язового волокна виникає під впливом імпульсів, що приходять по нервовому волокну. Аксони мотонейронів, розгалужуючись на своєму кінці, підходять до багатьом м’язових волокон (Рис. 37). Мотонейрон разом із усіма м’язовими волокнами, з якими він утворює синаптичне утворення, називають нейромоторною чи руховою одиницею. М’яз може включати декілька сотень нейромоторных одиниць. У відповідь на імпульси, що приходять від мотонейрона, скорочуються всі м’язові волокна, що входять у нейромоторну одиницю. Тому її називають функціональною одиницею діяльності нервово-м’язового апарату.
Особливості перебігу збудження. Мембранний потенціал спокою м’язового волокна дорівнює – 90 мв. Для виникнення збудження, що поширюється, необхідно щоб відбулася деполяризація до – 50 мв. По досягненні цієї величини з’являється ПД, амплітуда якого дорівнює 130 мв. Механізм підтримки в спокої різниці потенціалів на мембрані, так само як і механізм виникнення ПД для м’язового волокна, в основному такий же, як і для нервового.
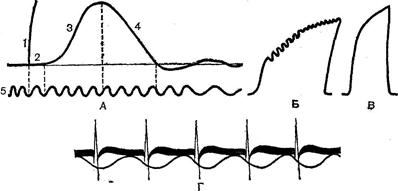
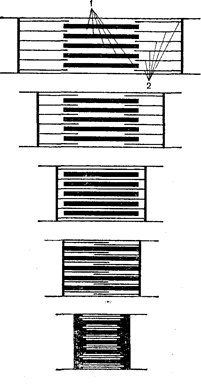
Рис. 38. Збудження і скорочення поперековопосмугованого м’язу:
А – аналіз одиночного скорочення поперековопосмугованого м’язу: 1 – оцінка подразнення; 2 – латентний період; 3 – скорочення м’язу; 4 – розслаблення м’язу; 5 – відмітка часу; Б – зубцюватий тетанус; В – суцільний тетанус;
Г – потенціали дії (верхня крива) і хвилі скорочення (нижня крива) поперековопосмугованого м’язу.
Імпульс поширюється по поверхневій мембрані м’язового волокна зі швидкістю 5–12 м/сек (у людини). Тривалість ПД складає близько 2 мсек. У глибину м’язового волокна збудження поширюється по поперечних трубочках і мембранах сарко-плазматичного ретикулюма. Вихід Са++ з нього починається при деполяризації мембран поперечних трубочок до 60 мв і триває протягом усього часу деполяризації. Припиняється вихід Са++, коли величина мембранного потенціалу знову стане (за абсолютним значенням) більше 60 мв.
Вихід Са++ супроводжується виникненням скорочення м’язу. Час між початком виникнення ПД і скороченням м’язу визначає латентний період скорочення. У цей період здійснюється передача збудження в глибину волокна. Деполяризація мембран поперечних трубочок стає граничною для виходу Са++ через 0,8 мсек (половина латентного періоду). З цього періоду починається активація скорочувального апарату, що забезпечує ковзання ниток.
Поодиноке скорочення м’язу біжить у вигляді хвилі, де можна розрізнити фазу підйому, зв’язану зі скороченням, і фазу спуску, названу розслабленням (Рис. 38, А). Тривалість фази скорочення і розслаблення неоднакова в різних м’язах і в різних умовах. Розслаблення, як правило, триває більше скорочення. У природних умовах до м’язу з центральної нервової системи надходять завжди серії імпульсів, на які вона реагує тетанічним скороченням. ПД м’язового волокна, а отже, і його рефрактерність приходяться на латентний період скорочення (Рис. 38, Г). Це є передумовою накладування хвиль скорочення одної на одну, що називають суперпозицією. Вона виникає у випадку, якщо кожен наступний імпульс приходить у момент, коли не закінчилася попередня хвиля скорочення. Оскільки волокно в цей період уже вийшло зі стану рефрактерності, можливе виникнення нового ПД, що викликає нову хвилю скорочення. Остання, сумуючись з попередньою, збільшує час скорочення м’язу. Якщо кожна нова хвиля скорочення виникає в момент, коли м’яз уже почав розслаблюватися під впливом попереднього подразнення, виникає зубцюватий тетанус (Рис. 38,Б). При меншому інтервалі між подразненнями і, отже, більшій їх частоті утвориться суцільний, чи гладкий,тетанус(Рис. 38,В). Для утворення гладкого тетанусу необхідно, щоб кожна нова хвиля скорочення виникала до початку м’язового розслаблення. У м’язових волокнах теплокровних тривалість наростання інтенсивності скорочення м’язу до його максимальної точки складає близько 0,02 сек. Звідси випливає, що для одержання суцільного тетанусу, коли відсутні інтервали між окремими хвилями скорочення, необхідно, щоб частота подразнення була не менш 50 імп/сек. Дійсно, якщо одне скорочення триває 0,02 сек, то в 1 сек їх повинно бути 50. Для виникнення 50 скорочень необхідно, щоб по руховому волокну пройшло таке ж число подразнюючих імпульсів. Тим часом відомо, що мотонейрони посилають до м’язу від 5 до 35 імп/сек і лише при максимальних скороченнях число їх може досягати 50 імп/сек. Виходить, злитий тетанус у природних умовах діяльності організму не може бути забезпечений частотою, що задається мотонейронами. Виявилося, що дійсно окремі волокна м’язу при її природній стимуляції з нерву відповідають на кожен імпульс одиночним скороченням. Злитий же тетанус виходить за рахунок сумації скорочень окремих м’язових волокон. Звичайно м’язові волокна скорочуються асинхронно відповідно неодночасному надходженню імпульсів від різних мотонейронов. Це також сприяє утворенню і підтримці злитого тетанусу.
Сила скорочення м’язів. Сила скорочення одного і того ж м’язу залежить від кількості нейромоторных одиниць, що беруть участь у цьому скороченні. Чим більше їх число, тим воно інтенсивніше. Сила залежить від частоти подразнення. До відомої межі збільшення частоти супроводжується зростанням сили м’язового скорочення. Це пов’язано з тим, що при збільшенні частоти велика кількість м’язових волокон включається в реакцію. Максимальна напруга, що може розвинути м’яз, визначається числом утворюючих її волокон: чим воно більше, тим більше сила м’язів. Тому перисті м’язи, у яких велике число волокон, відрізняються великою силою.
Прояв сили залежить від особливостей прикріплення м’яза до кісток. М’язи з більшою площею опори мають великі можливості для прояву сили. Важливо також і місце прикладення сили м’язів. Кістки разом з м’язами, що прикріплюються до них, є важелями, і, чим далі від точки опори важеля і ближче до точки прикладення сили ваги прикріплюється м’яз, тим більшу силу він може розвинути.
Залежність м’язової сили від цих факторів чітко виявляється в діяльності м’язів верхньої і нижньої кінцівки. Верхня кінцівка призначена для виконання різноманітних точних і швидких рухів. Функція нижніх кінцівок вимагає великої сили їх м’язів. Відповідно цим функціональним завданням неоднозначний характер прикріплення цих м’язів. Так, дельтоподібний м’яз, розташований в ділянці плечового суглоба, має невелику поверхню опори і прикріплюється на плечевій кісті поблизу від місця опори важеля. Навпаки, для м’язів нижніх кінцівок площа опори велика і точка додатка сили знаходиться далеко від точки опори. У сідничного м’язу площа опори в 23 рази більше, ніж у дельтоподібного, а площа прикріплення – у 4,5 рази.
Між силою м’язу і можливою найбільшою величиною його укорочення немає прямо пропорційної залежності. Максимальне укорочення м’язу, а виходить, і інтенсивність викликаного цим укороченням руху в тому чи іншому суглобі залежить від довжини м’язового волокна. Вона найбільша в м’язах з рівнобіжним розташуванням волокон, у той час як більшою силою володіють перисті м’язи. Попередньо розтягнутий м’яз при скороченні коротшає на велику величину.
Швидкість скорочення різних м’язових волокон неоднакова. У залежності від властивостей нейромоторних одиниць розрізняють декілька м’язових волокон: фазні, тонічні і перехідні.
Фазні нейромоторные одиниці беруть участь у здійсненні швидких рухів. Їх мотонейрони відносяться до групи α-мотонейронів, аксони яких входять у групу волокон Аα. М’язові волокна, що входять у ці рухові одиниці, відрізняються малою тривалістю ПД, рефрактерності хвилі скорочення і відносно великою лабільністю. Виділяють швидкі і повільні фазні волокна. У повільних фазних волокнах тривалість ПД і швидкість проведення збудження в 2 рази менше, а тривалість хвилі скорочення в 5 разів більше, ніж у швидких. У повільних фазних волокнах міститься багато міоглобіну (м’язового гемоглобіну). Тому м’язи, що містять велику кількість цих волокон, мають червоний колір і називаються «червоними» м’язами. Ті м’язи, що у великій кількості містять швидкі волокна, називають «білими». У кожному м’язі містяться і ті й інші волокна, і їх співвідношенням визначається швидкість його скорочення в цілому.
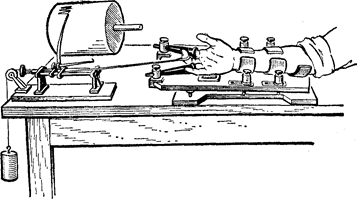 |
Рис. 39. Єргограф Моссо.
Тонічні нейромоторні одиниці здійснюють повільні реакції, їх діяльність забезпечує тонус м’язів. У тонічних волокнах м’язу виникає лише місцеве збудження в ділянці синапсу. Однак, деполяризується все м’язове волокно, тому що нервове волокно тонічної нейромоторной одиниці сильно розгалужується, утворюючи багато синапсів. Скорочення в тонічних волокнах розвивається дуже повільно, тривалість його велика. У деяких м’язах разом з фазними розташовані і тонічні волокна.
Перехідні нейромоторні одиниці функціонують і як фазні, і як тонічні: мала частота імпульсів викликає тонічні рефлекторні реакції, збільшення її супроводжується виникненням тетанічного скорочення.
Динамічна робота, пов’язана з переміщенням, обумовлюється діяльністю фазних і перехідних волокон, а статична, спрямована на підтримку визначеного положення тіла, залежить від функціонування тонічних і перехідних нейромоторних одиниць.
Робота м’язу. М’яз, скорочуючись, робить роботу. Величина її дорівнює добутку вантажу на підняту висоту. Звідси випливає, що максимальна робота, виконувана при одиночному скороченні м’язу, залежить від її сили (чим більше сила, тим більший вантаж може бути піднятийі) і ступені укорочення м’яза.
У процесі природної діяльності людини величина роботи, виконуваною тим чи іншим м’язом, у значній мірі залежить від здатності її довгостроково знаходитися в скороченому стані, тобто від ступеня витривалості м’язів. Розрізняють витривалість до статичних і динамічних зусиль. Витривалість до статичних зусиль визначається часом, протягом якого утримується величина заданого зусилля. Вона різна для різних м’язів. Найменшою витривалістю характеризується триголовий м’яз плеча (1 хв при зусиллі, рівному 50% від максимального), найбільшою–литковий м’яз (7 хв).
Для оцінки витривалості до динамічної роботи здавна користалися ергографом Моссо, за допомогою якого реєструються рухи вказівного чи середнього пальця, здійснювані в різних ритмах і при різній величині навантаження. Ці дослідження показали, що витривалість до динамічної роботи залежить не тільки від величини вантажу, що піднімається, але і від темпу роботи. Робота буває найбільшою при якійсь середній величині вантажу і частоті рухів. Витривалість до динамічної і статичної роботи можна збільшувати шляхом тренування. При тривалій як динамічній, так і статичній роботі настає стомлення м’язів. В експерименті воно виявляється в зменшенні амплітуди скорочення, збільшенні тривалості періодів одиночного скорочення, появі контрактури – залишкового скорочення внаслідок неповного розслаблення.
У природних умовах діяльності людини м’язове стомлення є наслідком процесів, що розвиваються у всіх ланках рефлекторної дуги.
РОЗДІЛ VІ ВІКОВІ ОСОБЛИВОСТІ
ОПОРНО-РУХОВОГО АПАРАТУ
ВІКОВІ ОСОБЛИВОСТІ КІСТЯКА
Ріст і розвиток кісток. В ембріональному періоді розвитку кістяк закладається як сполучнотканинне утворення. У деяких кістках безпосередньо в сполучнотканинному скелеті з’являються вогнища окостеніння, тобто кістка у своєму розвитку минає хрящову стадію. Такі кістки називають первинними (кістки черепа). Для більшості кісток характерне заміщення сполучної тканини хрящовою, після чого хрящ руйнується і замість нього утвориться кісткова тканина. Так формуються вторинні кістки.
Окостеніння відбувається двома шляхами: енхондральне окостеніння, коли вогнища окостеніння з’являються усередині хряща, і періхондральне, що починається з його поверхні.
У колагенових волокнах кісткової тканини, що формується, (у визначених їхніх ділянках) містяться активні центри кристалізації, що мають реакційноздатні групи. Припускають, що процес звапнення починається з взаємодії амінокислоти лізина, що входить у реакційну групу колагену, з іонами фосфату. На перших стадіях мінералізації кристали неорганічних солей не орієнтовані щодо осей колагенових фибрил. Але в міру мінералізації кристали, що утворяться, орієнтуються своїми довгими осями паралельно осям тих колагенових фибрил, з якими вони пов’язані. В епіфізах, у коротких кістках, у відростках кісток окостеніння здійснюється по енхондральному типу, а в діафизах – по періхондральному. Окостеніння починається із середньої частини діафізу, де утвориться завдяки діяльності остебластів кісткова манжетка. Кісткова манжетка росте в напрямку до епіфізів. Разом з тим відбувається збільшення її товщини за рахунок утворення все нових і нових шарів кісткової тканини. Одночасно усередині відбувається розсмоктування хрящової і кісткової тканин, при цьому формується косткомозкова порожнина. Таким чином, зовні відбувається все нові нашарування шарів кісткової тканини, а зсередини руйнування залишків хрящової і кісткової тканин. За рахунок цього кістка росте в товщину. На визначеному етапі ембріонального розвитку з’являються вогнища окостеніння в епіфізах. Однак тривалий час на межі діафізу й епіфізу зберігається хрящова зона – пластинка росту, що обумовлює здатність росту кісток у довжину.
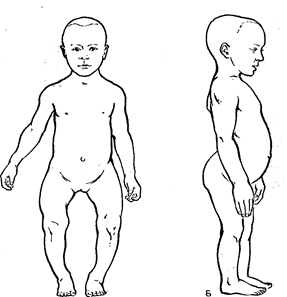
Рис. 40. Зміни кістяка при рахіті:
А – скривлення ніг; Б – деформація черепа, хребта, грудної клітки.
Для здійснення складного процесу формування кісток необхідно повноцінне як у якісному, так і в кількісному відношенні харчування. Їжа дітей повинна містити в достатній кількості солі Р и Са, без яких неможливий процес звапнення, а також необхідна кількість вітамінів. Так, недолік вітаміну А викликає звуження судин окістя і пов’язаний з цим розлад харчування кісткової тканини, що формується, у результаті чого кістка перестає рости. При недоліку вітаміну С не формуються кісткові пластинки. При недоліку вітаміну D порушується обмін фосфору і кальцію. Виникає захворювання – рахіт,–, що виявляється в порушенні процесу формування кісткової тканини. Це захворювання характеризується пом’якшенням кісткової тканини і деформацією внаслідок цього кісток, а також посиленим розростанням тканини, що відрізняється від кісткової своєю структурою і хімічним складом (Рис. 40).
Вікові особливості структури кісток. Окостеніння починається у внутрішньоутробному періоді розвитку, коли з’являються первинні ядраокостеніння. Значно більше число ядер окостеніння виникає після народження дитини. Ці ядра називаються вторинними. Усього за час розвитку в кістяку людини формується 806 ядер окостеніння.
Тільки в черепі майже всі ядра окостеніння з’являються у внутрішньоутробному періоді розвитку. В усіх же інших частинах кістяка число вторинних ядер більше числа первинних. У дорослого число кісток значно менше, ніж у підлітка 14 років: у дорослого–206, у 14 років–356. Звідси випливає, що і після 14 років продовжується зрощення кісток.
Кістка немовляти характеризується великою кількістю хрящової тканини, великою товщиною окістя, багатою судинною мережею, неправильним розташуванням гаверсовых каналів. Щойно сформована кісткова тканина багата водою. Неорганічна речовина кістки складає лише половину її ваги. Усе це робить кістку менш щільною, порозною, більш пружною, еластичною і гнучкою, така кіста легко змінює свою форму.
Кожен відділ кістяка має свої особливості розвитку.
Вікові особливості кістяка черепа. Череп починає диференціюватися на 2-му місяці внутрішньоутробного життя. Кістки черепа розвиваються і первинним, і вторинним шляхом. До моменту народження ядра окостеніння розміщені у всіх кістках черепа, але їх розростання і зрощення відбувається в постнатальному періоді. У новонародженого обсяг мозкового черепа в 8 разів більше лицьового, а у дорослого тільки в в 2–2,5 рази. У 2 роки відношення обличчя/череп дорівнює 1:6, у 5 років– 1:4, у 10 років – 1:3 (Рис. 41). Менша величина лицьового черепа в немовлят залежить від недорозвинення лицьових, головним чином щелепних, кісток. З ростом зубів ці співвідношення наближаються до співвідношення їх у дорослого.
У немовляти між кістками черепа є простори розміром близько 3 мм, заповнені сполучною тканиною. Їх називають швами. У процесі постнатального розвитку ширина швів зменшується, так що сполучнотканинний прошарок стає ледь помітним. Після30 років відбувається окостеніння швів.
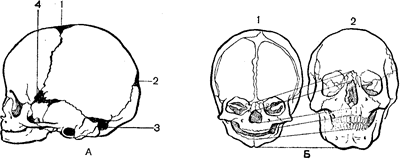
Рис. 41. Особливості черепа немовляти:
А – розташування тім’ячок: 1 – лобове; 2 – потиличне; 3 – заднє бічне; 4 – переднє бічне; Б – співвідношення між лицьовою і мозковою частинами черепа в новонароджених і дорослих: 1 – у немовляти; 2 – у дорослого.
Кути кісток черепа не костеніють до моменту народження, і місця їх з’єднання також заповнює сполучна тканина. Ці ділянки називають тім’ячками(Рис. 41, А). Розрізняють переднє, заднє і бічне тім’ячка Переднє, лобове тім’ячкорозташоване між лобовою і тім’яною кістками, його розмір складає 2,5–5 см. Воно прогресивно зменшується до 6 місяців постнатального розвитку і цілком закривається до 1,5–2 років. Заднє, потиличне тім’ячкознаходиться між потиличною і тім’яною кістками, воно має розмір до 1 см. Звично воно уже закрите до моменту народження, але іноді зберігається до 4–8 тижнів. Бічне переднє тім’ячкоміститься в місці сходження лобової, тім’яної, основної і скроневої кісток, а бічне заднє – між потиличною і скроневою кістками. Їх закриття відбувається або у внутрішньоутробному періоді розвитку, або в перші тижні після народження. При рахіті закриття тім’ячоквідбувається в більш пізній термін. Тривале збереження тім’ячок вважається одним із симптомів цього захворювання. Наявність до моменту народження джерелець і швів має велике значення, тому що дозволяє зміщатися кістам черепа дитини при його народженні, полегшуючи тим самим проходження через родові шляхи матері.
Потилична кістка в немовлят складається з чотирьох незрослих кісток, скронева – із трьох, нижня щелепа –іздвох половин, лобова – із двох, у клиноподібній кістці не зрощені передня і задня частини її тіла, а також крила з тілом. На першому році життя зростаються великі крила з тілом клиноподібної кістки, зрощення передніх і задніх ділянок її тіла відбувається тільки в 13 років. Половинки нижньої щелепи зростаються до 2 років. Зрощення окремих частин скроневої кісти відбувається в 2–3 роки, потиличної у 4–5 років. Зрощення двох половин лобової кістки закінчується до кінця 3-го року життя, шов між ними зникає в 7–8 років.
Пазухи в кістках черепа формуються в основному після народження дитини. У немовляти є тільки зачаток верхньощелепної, чи гайморової, порожнини. Формування пазух закінчується тільки в зрілому віці.
У немовляти кістки черепа дуже тонкі, їх товщина в 8 разів менше, ніж у дорослого. Однак завдяки інтенсивному процесу кісткоутворення вже на першому році життя товщина стінок збільшується в 3 рази.
Досить швидко змінюється обсяг черепа: у новонародженого він складає 1/з, у 6 місяців – 1/2, а до 2 років – 2/з обсягу черепа дорослого. З 10–12 років величина його міняється дуже мало.
Вікові особливості кістяка тулуба. Хребці, що формують хребетний стовп, розвиваються як вторинні кістки, тобто вони проходять хрящову стадію. Ядра окостеніння в них з’являються на 2-му місяці внутрішньоутробного розвитку. Процес окостеніння хребетного стовпа відбувається в строго визначеному порядку. Вогнища окостеніння спочатку з’являються в грудних хребцях, і потім окостеніння поширюється в напрямку до шийного відділу і куприкового.
На 40–50-ій добі внутрішньоутробного розвитку ядро окостеніння з’являється в тілі 12-го грудного хребця, до кінця 4-го місяця тіла всіх грудних хребців, шийних, поперекових і двох перших крижових мають ядра окостеніння. У цей же період з’являються ядра окостеніння в інших хребцях. Злиття ядер окостеніння правої і лівої половин дуг хребців відбувається тільки після народження. Хребет немовляти відкритий позаду по лінії всіх дуг хребців. Тільки до 7 років усі дуги виявляються закритими. Виключенням може бути лише дуга першого крижового хребця. Іноді вона закривається пізніше. Передня дуга атланта може залишатися відкритою до 9 років.
У 8–11 років з’являються ядра окостеніння в епіфізарних хрящових дисках, що обмежує хребці зверху і знизу. З 15 до 24 років відбувається зрощення кісткових епіфізарних дисків з тілом хребця. Раніш усього це відбувається в грудному відділі хребта, потім у шийному і поперековому. Повне зрощення відростків з тілом хребця здійснюється у віці 18–24 років.
Тіла хребців у немовлят сплющені так, що їх поперечний діаметр більше подовжнього і співвідношення між діаметрами дорівнює 5:3. У період статевого дозрівання це співвідношення стає рівним 4:3, a y дорослого – 3:3. У цілому за весь період розвитку довжина хребта збільшується в 3,5 рази. Перші 2 роки ріст хребта дуже інтенсивний,потім він сповільнюється і стає більш інтенсивним в період статевого дозрівання.
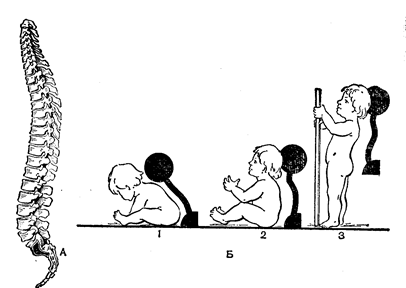
Рис.42. Вигини хребта.
Хребетний стовп немовляти має тільки невеликий крижовий вигин (Рис. 42). Першим з’являється шийний вигин у віці 2,5–3 місяців, коли дитина починає тримати голівку. Вигин, спрямований опуклістю вперед, називають лордозом. Отже, першим з’являється шийний лордоз. У віці близько 6 місяців, коли дитина починає сидіти, виникає вигин у грудному відділі, спрямований опуклістю назад. Такі вигини, спрямовані опуклістю назад називають кіфозами. До моменту початку ходьби формується поперековий вигин. Це супроводжується зміною положення центра ваги, що запобігає падінню тіла при переході до вертикального положення. Таким чином, до року існують уже усі вигини хребта. Спочатку утворилися вигини не фіксовані і зникають при розслабленні мускулатури. Фіксація вигинів у шийному і грудному відділах хребта відбувається в 6–7 років, а в поперековому –до 12 років.
Окостеніння грудини відбувається вторинним способом, причому перші ядра окостеніння з’являються в рукоятці і тілі її ще у внутрішньоутробному періоді розвитку. У мечоподібному відростку ядро окостеніння виникає лише в 6–12 років.
Повне зрощення всіх кісткових ділянок грудини здійснюється після 25 років.
Окостеніння хрящових ребер починається на 6–8-й тиждень внутрішньоутробного розвитку. Раніше всього з’являються ядра в середніх ребрах. Вторинні ядра виникають у 8–11 років. Злиття кісткових частин ребра відбувається у віці 18–19 років, а голівки і тіла ребра – у 20–25 років.
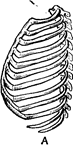
У немовлят грудна клітина має форму чи дзвону груші. Верхня частина грудної клітини вузька, нижня через високе розташування внутрішніх органів розширена, передне-задний діаметр її більше поперечного (Рис. 43). З розвитком легень, що починають займати великий простір, верхні ребра, що розташовувалися косо, починають займати горизонтальне положення. У зв’язку з цим грудна клітина приймає бочкоподібну форму. Верхній край грудини в немовляти знаходиться на рівні першого грудного хребця. Зігнутість ребер невелика. Кут між ребрами і хребтом, так само як і між ребрами і грудиною, великий. Так, реберно-хребетний кут у немовляти складає 82°, а в 3 роки – 62°. Форма грудної клітини в цей період відповідає фазі максимального вдиху. Звідси зрозуміло, що подих у цьому віці здійснюється в основному за рахунок діафрагми. До 3– 4 років верхній край грудини опускається до рівня 3–4-го грудного хребця (як у дорослих). Разом із грудиною опускаються ребра, збільшується їх зігнутість, зменшуються ребернохребтовий кут і кут між ребрами і грудиною. Це приводить до усе більшої залежності акту дихання від зміни обсягу грудної клітини. Ця залежність уже чітко виявляється в 3-літньої дитини.
|
|
Рис. 43. Різні форми грудної клітини:
А – у новонароджених, Б – у дорослих.
Форму дорослого грудна клітина здобуває до 12–13 років.
Вікові особливості кістяка кінцівок. Всі кістки пояса верхніх кінцівок, за винятком ключиці, проходять хрящову стадію. У ключиці передхрящова тканина відразу заміщається кістковою. Процес окостеніння, що почався в ній на 6-й тиждень внутрішньоутробного розвитку, майже цілком закінчується до моменту народження. Лише грудинний кінець ключиці не має ядра окостеніння. Воно з’являється тільки до 16–22 років, а зрощення його з тілом відбувається до 25 років.
У більшості кісток вільних верхніх кінцівок первинні ядра окостеніння виникають протягом 2–3 місяців ембріонального розвитку. У кістках зап’ястя вони з’являються після народження: у голівчатій і гачкуватій – на 4–5-му місяці, а в інших – у період від 2 до 11 років. Зрощення первинних і вторинних ядер окостеніння в кістках пояса закінчується до 16– 25 років.
Майже у всіх кістках пояса нижніх кінцівок первинні ядра окостеніння з’являються також в ембріональному періоді розвитку. Лише в кістках передплесно(човнуватої, кубоподібної і клиноподібної) вони утворяться в період від 3-х місяців після народження до 5 років.
Таз у немовляти має форму лійки. Його передньо-задній розмір більше поперечного. Нижній отвір тазу дуже малий. Площина входу розташована значно більш вертикально, ніж у дорослого. Таз немовляти складається з окремих, незрослих кісток. Ядра окостеніння в здухвинній, сідничній і лобковій кістках з’являються в період від 3,5 до 4,5 місяців внутрішньоутробного розвитку. З 12 до 19 років з’являються вторинні ядра окостеніння. Зрощення всіх трьох кісток таза відбувається в 14–16 років, а вторинні ядра з’єднуються з раніше сформованими і кістами таза, що зрослися, тільки до 25 років.
У постнатальному періоді відбувається зміна форми і розміру таза під впливом усіляких факторів: під впливом тиску, що робиться масою тіла й органами черевної порожнини, під впливом м’язів, у результаті тиску голівки стегнової кісти, під впливом статевих гормонів і т.д. У результаті цих різноманітних впливів збільшується передньо-задній діаметр таза (з 2,7 см у немовляти до 8,5 сму 6 років і 9,5 см у 12 років), зростає його поперечний розмір, що у 13–14 років стає таким же, як і в дорослих. Площина тазу в поперечному діаметрі стає в цьому віці овальною.
Після 9 років відзначається різниця у формі тазу в хлопчиків і дівчаток: у хлопчиків таз більш високий і більш вузький, ніж у дівчаток.
ВІКОВІ ОСОБЛИВОСТІ КІСТЯКОВОЇ МУСКУЛАТУРИ
Зміни макро - і мікроструктури кістякових м’язів з віком. Формування кістякових м’язів відбувається на дуже ранніх етапах розвитку. На 8-й тиждень внутрішньоутробного розвитку помітні уже всі м’язи, а до 10-го тижня розвиваються їх сухожилля. Зв’язок первинної закладки м’язів з відповідними нервами виявляється вже на 2-му місяці розвитку. Однак рухові нервові закінчення вперше з’являються лише на 4-му місяці внутрішньоутробного розвитку.
Дозрівання м’язових волокон пов’язане із збільшенням кількості міофибрил, появою поперекової посмугованості, збільшенням числа ядер. Воно здійснюється в різних м’язових волокнах з неоднаковою швидкістю. Раніше всього диференціюються волокна м’язів язика, губ, міжреберних м’язів, м’язів спини і діафрагми. Потім – м’язи верхньої кінцівки й в останню чергу – м’язи нижньої кінцівки.
У новонароджених маса м’язів складає 23,3% (у дорослих–44,2%) від маси всього тіла. Сухожильна частина м’язу розвинута слабко і складає меншу, ніж у дорослих, частину від усієї довжини м’язу; фасції і сухожилля широких м’язів дуже тонкі, неміцні, легко від них відокремлюються. Сполучна гканина, що утворює внутрім’язові перетинки, відрізняється від сполучної тканини м’язів дорослого великою кількістю клітин і меншим числом волокон. Поперереково посмуговані волокна характеризуються дуже великим числом ядер, що мають овальну форму. Подовжній діаметр відноситься до поперечного, як 2:1. Різні м’язові волокна у новонароджених мало відрізняються в своєму діаметрі.
Маса м’язів верхніх кінцівок стосовно маси м’язів усього тіла зростає від народження до 23–25 років, коли закінчується онтогенетическое дозрівання м’язів, усього лише на 2%. Отже, до моменту народження вони вже мають досить велику масу, і подальше збільшення її йшло в повній відповідності зі збільшенням маси всього тіла. У той же час маса м’язів нижніх кінцівок стосовно маси тіла збільшується за весь період розвитку більш ніж на 16%. У м’язах верхніх кінцівок особливо різко збільшується в дошкільному і молодшому шкільному віці маса тих з них, що викликають рухи пальців. Маса м’язів – розгиначів збільшується інтенсивніше, ніж згиначів, тому що до моменту народження згиначі, що обумовлюють у період внутрішньоутробного розвитку характерну позу плоду, повинні бути вже значно розвинуті. Розгиначі, що забезпечують вертикальне положення тіла, інтенсивно дозрівають після народження дитини.
М’язи, що обумовлюють великий розмах руху, інтенсивно ростуть у довжину, а м’язи, функція яких вимагає скорочень великої сили, збільшуються в діаметрі. Розвиток їх характеризується ростом ступеня перистості.
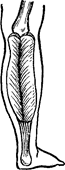
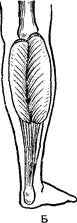
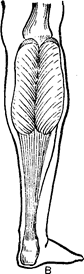
З віком відбувається збільшення довжини сухожилля. Так, у новонародженого довжина сухожилля литкового м’язу стосовно довжини всього м’язу складає 0,25, а в 1,5 року–0,5 (Рис. 44). До 12–14 років відношення довжини сухожилля і черевця м’яза стає таким же, як і в дорослих.
|
|
|
Рис. 44. Вікові зміни співвідношень
черевця литкового м’язу і його сухожильного відділу:
А – немовляти; Б – дитини 1 року 6 місяців; В – дитини 6–7 років.
Ріст м’язів у довжину може продовжуватися до 23–25 років. Він здійснюється за рахунок зони росту, розташованої на межі м’язової і сухожильної частин. У зоні росту є скупчення ядер, число яких з віком зменшується, причому особливо значно після 7 років. До 15–18 років зона росту зменшується в 3 рази. До 15–16 років закінчується формування сарколеми, коли її волокна здобувають визначену орієнтацію; вони спрямовані перпендикулярно до подовжньої осі м’язового волокна. Розвиток у постнатальному онтогенезі сполучної тканини м’язів характеризується зменшенням числа клітин, що приходяться на одиницю площі, і збільшенням числа волокон. Так, до 3–4 років число клітин сполучної тканини, що приходяться на одиницю площі, складає 25–30 (у немовляти–50–60). Навіть у 13–15 років число клітин усе ще в 2 рази більше, ніж у дорослих.
Діаметр м’язових волокон збільшується до 35 років. Зміна діаметра кожного окремого м’язового волокна і загального числа їх викликає наростання поперечника м’язів у процесі росту і розвитку організму. Різниця в діаметрі волокон одного і того ж м’язу відзначається уже в ембріональному періоді розвитку. Так, у двоголовому м’язі плеча на 5–6-м місяці внутрішньоутробного розвитку зустрічаються волокна діаметром і 2,5, і 12,5 мкм. Але особливо великою ця різниця стає після народження. Наприклад, товщина волокон великого сідничного м’язу і прямого м’яза ока в немовлят однакова, а в дорослих діаметр волокон одного м’яза більше діаметра іншого в 5 разів. Діаметр одних волокон збільшується в 5–9 разів (двоголовий м’яз плеча), інших у 4–5 разів (згинач пальців і кисті).
Найбільш інтенсивний ріст волокон відзначається в період статевого дозрівання. Так, у перші 6 місяців життя діаметр волокон двоголового м’яза плеча складає 17–19 мкм, у 2–3 роки – 20 – 22 мкм, у 9 –12 років – 20 – 25 мкм, а в дорослих – 41–5 8 мкм.
З віком росте число міофибрил за рахунок їх подовжнього розщеплення. У немовляти в кожнім м’язовому волокні міститься 50–120 міофібрил, у 1,5 року їх число стає в 2 рази більшим, у 3–4 роки воно збільшується в 5–6 разів, до 7 років у 15–20 разів.
У процесі дозрівання кістякової мускулатури міняється форма і кількість ядер, що приходяться на одиницю площі. До 3 років кількість ядер зменшується в порівнянні з їх числом у немовлят приблизно в 2 рази, до 5 років у 2,5 рази і до 7 років– у 3 – 4 рази. Ядра з овальних приймають паличкоподібну форму, у якій довжина перевищує поперечник у 4 рази. В міру розвитку м’язів ядра усе більш переміщаються до периферії. Розташування ядер і їх число, характерне для дорослих, відзначається вже у віці 7–10 років.
Хімічний склад м’язів з віком також міняється. М’язи дітей містять більше води, вони багаті нуклеопротеїдами. В міру росту відбувається неухильне наростання актоміозину й АТФ, креатинфосфорної кислоти, м’язового гемоглобіну (міоглобіну). У зв’язку з тим що міоглобін є джерелом кисню, збільшення його кількості сприяє удосконалюванню скорочувальної функції м’язу.
Розвиток рухових нервових закінчень відбувається особливо інтенсивно в перші місяці після народження. Але тільки до 11–13 років цілком закінчується їх структурне оформлення.
Вікові особливості функції кістякових м’язів. Морфологічний субстрат скорочувального акту формується на ранніх стадіях онтогенезу. Кістякові м’язи вже до семи тижнів внутрішньоутробного розвитку здатні скорочуватися, а до 12 тижнів виникають елементарні рефлекторні рухи у відповідь на механічні подразнення. У 12 тижнів плід може стискати кисть у кулак.
Скорочення в ембріональний період розвитку носить тонічний характер: воно повільно наростає і потім довгостроково утримується на одному рівні. Такий характер реакції обумовлюється вмістом у м’язі великої кількості волокон типу «червоних».
Процес збудження м’язового волокна, що передує його скороченню, здобуває характеристики, властиві м’язам дорослих, не відразу. У дітей мембранний потенціал і ПД відрізняються по своих характеристиках від таких у дорослих.
Збільшення змісту іонів внутрішньоклітинного калію відбувається до 9 років. Оскільки іони К визначають величину мембранного потенціалу спокою, значення її протягом перших 9 років життя в дітей менше, ніж у дорослих.
Повне функціональне дозрівання м’язів відбувається на пізніх етапах розвитку. Електрозбудливість м’язів дітей в ембріональний період і в перші місяці постнатального розвитку менше, ніж у дорослих, тобто поріг подразнення більш високий. Хронаксія кістякових м’язів тим більше, чим менше вік дитини. Величина хронаксії м’язів немовлят перевершує хронаксію м’язів дорослих у 1,5–10 разів. Величина хронаксії, характерна для м’язів дорослих, установлюється до 10–17 років. Для різних м’язів ці терміни виявляються неоднаковими. Так, хронаксія двоголового м’язу в підлітків 14–19 років більше, ніж у дорослих, а хронаксія загального згинача пальців дорівнює хронаксії дорослих.
Змінюється з віком і відношення хронаксії згиначів до хронаксії розгиначів. У дорослого воно дорівнює 1:2. У новонароджені величини хронаксії згиначів і розгиначів відрізняються дуже мало, а іноді хронаксія згиначів виявляється навіть більше хронаксії розгиначів. Співвідношення цих величин, характерне для дорослих, установлюється після 10 років.
Велика величина хронаксії в дітей у порівнянні з величиною її в дорослих служить непрямим свідченням меншої швидкості й інтенсивності протікання в них процесів, що викликають деполяризацію мембрани м’язового волокна до критичного рівня.
Чим менше вік дітей, тим менше величина лабильності їх м’язових волокон. Звідси випливає, що тривалість процесів реполяризації з віком збільшується.
Дозрівання волокон одного і того ж м’язу відбувається неодночасно. Частина волокон має лабільність, характерну для м’язів дорослих уже у віці 10–13 років, а лабільність інших волокон продовжує збільшуватися аж до 14–15 років.
Прямі визначення тривалості ПД в окремих волокнах м’язу показали, що величина його у віці 14 років складає 5,7 мсек проти 2 мсек у дорослих. З віком збільшується м’язова сила за рахунок росту діаметра м’язових волокон і їх числа Для різних м’язів найбільша величина їх сили відзначається в різному віці. Більшість м’язів верхньої кінцівки і шиї досягає найбільшої сили до 20–30 років. У той же час сила м’язів, що розгинають тулуб, стає максимальною до 16 років. Різниця між силами м’язів-згиначів і м’язів-розгиначів збільшується з віком.
Характеристикою функціонального дозрівання м’язів служить м’язова витривалість. У дошкільному і молодшому шкільному віці відбувається найбільше в порівнянні з іншими віками збільшення витривалості. Однак, навіть у 16–19 років її величина складає лише 85% від витривалості дорослого.
Вивчення характеру електричної активності м’яза при довільному максимальному його напруженні дозволило вважати, що менша здатність дітей зберігати максимальну величину статичного зусилля залежить від швидко розвиваючого в них процесу асинхронизації роботи м’язових волокон. Чим менше вік, тим менший час може зберігатися одночасне збудження волокон м’язу і тим швидше в ній настає стомлення.
Морфологічне і функціональне дозрівання опорно-рухового апарата залежить від дуже багатьох факторів: спадкоємних, умов життя, харчування, рухової активності. Правильно організовані заняття фізичною культурою і спортом сприяють формуванню як кісткової, так і м’язової системи. Під впливом цих занять збільшується діаметр м’язових волокон, росте їх число, більш досконалими стають координаційні відношения між м’язами-антагоністами. Однак варто пам’ятати, що м’язова діяльність викликає в дітей у порівнянні з дорослими значно більші зміни в діяльності всіх їх органів, що вимагає диференційованого підходу до організації всякого роду занять фізкультурою з дітьми різного віку.
РОЗДІЛ VІІ ЕНДОКРИННІ ЗАЛОЗИ
СТРУКТУРА І ФУНКЦІЇ ЕНДОКРИННИХ ЗАЛОЗ
Поняття про ендокринні залози і гормони. Ендокринними залозами називають органи, у яких утвориться секрет, що специфічно впливає на різні функції організму, регулюючи поряд з нервовою системою його діяльність. Секрет ендокринних залоз називають гормонами. Слово “гормон” походить від грецького hormao, що означає “збуджую”, “рухаю”. Ендокринні залози, на відміну від інших залоз організму, не мають вивідних проток, і їх секрет виводиться безпосередньо в кров чи лімфу. По цій, загальній для всіх ендокринних залоз ознаці вони одержали свою назву: грецьке слово “эндо” означає “всередині", а “крино” – відокремлюю. На підставі цієї ж ознаки ендокринні залози називають залозами внутрішньої секреції, підкреслюючи цим їх відмінність від залоз зовнішньої секреції (як, наприклад, шлункових), що мають вивідні протоки. До залоз внутрішньої секреції відносять гіпофіз, щитовидну, навколощитовидні, вилочкову. наднирники, епіфіз, підшлункову і статеві.
Гіпофіз, щитовидна залоза, навколощитовидна і наднирники мають тільки внутрішню секрецію. Підшлункова залоза і статеві залози характеризуються змішаною функцією: у них не тільки утворяться гормони, але і здійснюється секреція речовин, що не володіють гормональною активністю.
Гормони можуть синтезуватися в органах, що не відносяться до ендокринного. Так, наприклад, адреналін утворюється не тільки в наднирниках, але у всіх так званих хромофінних клітинах, які входять до складу різних органів. У клітинах слизової оболонки травного тракту, у плаценті утворюються речовини, що виділяються безпосередньо в кров і володіють здатністю регулювати ті чи інші функції організму (гастрин, секретин, естрогени та ін.).
Біологічні речовини, які утворюються не в ендокринних залозах, але мають здатність змінювати фізіологічні функції, виділяють в особливу групу – гормоноїдів.
Значення гормонів, гормони впливають на усі функції організму. Вони регулюють обмін речовин (білковий, жировий, вуглеводний, мінеральний, водний), тим самим сприяючи підтримці гомеостазу, під яким розуміють відносну сталість внутрішнього середовища організму. Вони впливають на ріст і формування органів, систем органів і всього організму в цілому. Під впливом гормонів здійснюється тканьове диференціювання, вони можуть спричинити пускову дію на органа ефектора чи змінити інтенсивність функціонування будь-якого органа. Залози внутрішньої секреції поряд з нервовою системою забезпечують пристосувальні реакції організму в умовах стресу, тобто напруги, викликаної зовнішніми впливами.
Методи вивчення залоз внутрішньої секреції. Методи вивчення залоз внутрішньої секреції дуже різноманітні.
1. Метод екстирпації, чи видалення, залози внутрішньої секреції з наступним спостереженням за зміною функцій організму.
2. Метод трансплантації, тобто пересадження залози. При цьому залоза пересаджується або в іншу ділянку тіла цієї ж особи, або в організм іншої особи.
3. Введення в організм витяжок ендокринних залоз чи їх гормонів і спостереження за змінами функцій організму, що відбуваються при цьому.
4. Визначення вмісту гормонів у крові і сечі.
5. Вивчення за допомогою радіоактивних ізотопів. В організм уводять гормон, що містить радіоактивний ізотоп (з міченим йодом, вуглецем, сіркою і т.д.). Через деякий проміжок часу визначають його вміст у різних тканинах. Цей метод дає можливість простежити, якими органами й у якій кількості захоплюється гормон, як і у виді яких продуктів відбувається його виділення.
6. Зіставлення даних спостережень за симптомами тих чи інших захворювань з даними посмертного анатомічного і гістологічного вивчення ендокринних залоз.
7. Метод радіоавтографії. Цей метод знайшов широке застосування у вивченні місця утворення і розподілу гормонів в організмі. Сутність його полягає в тім, що в організм уводиться радіоактивна речовина, що у наступному бере участь у побудові гормону. Метод радіоавтографії дозволяє точно локалізувати місце перебування гормону. Цим способом удалося виявити гормони навіть в окремих елементах клітини.
Успіхи у вивченні гормонів дозволили деякі з них штучно синтезувати. Так, штучно синтезуються деякі гормони гіпофіза, статевих залоз, підшлункової залози, наднирників (окситоцин, вазопресин, адреналін, інсулін, кортизон і ін.).
Структура гормонів. Дослідження хімічного складу гормонів показало, що вони представлені білками, поліпептидами, похідними амінокислот і ліпідами. Поряд з гормонами, що містять всього 8 амінокислот (вазопресин), є гормони, що містять 396 амінокислот (соматотропний гормон гіпофіза бика). Одні гормони представлені простими білками, інші – складними. Цілий ряд гормонів утвориться при обміні амінокислот.
Багато гормонів ліпідної природи є стеринами. До них відносяться статеві гормони і гормони кори наднирників.
Особливості дії гормонів. Для всіх гормонів характерна специфічність їхньої дії. Явища, що виникають при недостатності однієї з залоз, можуть зникнути тільки при лікуванні гормонами такої ж залози. Так, збудження вуглеводного обміну, що виникають при недостатності функції підшлункової залози, можна ліквідувати тільки гормоном цієї ж залози – інсуліном. Усі гормони можуть діяти на ті чи інші органи, розташовані на великій відстані від місця їх виділення. Гіпофіз розташований у порожнині черепа, а його гормон діє на різні орани, у тому числі і на статеві залози, розташовані в порожнині тазу. Гормони роблять ефект у дуже невеликих концентраціях, тобто їх біологічна активність висока. Наприклад, жіночого статевого гормону міститься в крові 0,6 мкг на 100 мл плазми.
Механізм дії гормонів. Дослідження останніх років привели до створення ряду гіпотез щодо механізму дії гормонів. Він неоднаковий для різних гормонів, неоднаковий при дії на різні субстрати й у різних представників світу тварин. Вважають, що гормони діють шляхом зміни фізичної структури ферментів, проникності клітинної мембрани і впливу на генетичний апарат клітини. Відповідно до першої гіпотези, гормони, приєднуючись до ферментів, змінюють
Читайте також:
- III мозковий шлуночок
- IV мозковий шлуночок
- Агресивний тип дивідендної політики включає метод стабільного приросту дивідендів і метод постійного коефіцієнта виплат.
- Асортиментний процес включає три основних етапи: концентрацію, кастомізацію і розсіювання.
- Аудит фінансово-господарської діяльності бюджетних установ включає три етапи.
- Банківська система в ринковій економіці є звичайно дворівневою і включає центральний банк (емісійний) і комерційні (депозитні) банки різних видів.
- Верховна Рада України обирає із свого складу Голову Верховної Ради України, Першого заступника і заступника Голови Верховної Ради України та відкликає їх з цих посад.
- Діяльність включає дії і операції як складові, які співвідносяться з потребами, мотивами і цілями.
- Дохідна частина бюджетної класифікації доходів бюджету України включає наступні статті доходів.
- Економічний цикл включає такі фази : кризу, депресію, пожвавлення та піднесення, яке знаходить остаточне відображення у новій кризі.
- Ідеї правової держави Б.Кістяківського і сучасність
- Із виступу Голови РНК УСРР X. Раковського на XII з’їзді РКП(б) 1923 р.
| <== попередня сторінка | | | наступна сторінка ==> |
| Хімічний склад, будова і з’єднання кісток. | | | Гормони аденогіпофіза, їх фізіологічне значення, механізм дії. |
|
Не знайшли потрібну інформацію? Скористайтесь пошуком google: |
© studopedia.com.ua При використанні або копіюванні матеріалів пряме посилання на сайт обов'язкове. |