РЕЗОЛЮЦІЯ: Громадського обговорення навчальної програми статевого виховання
ЧОМУ ФОНД ОЛЕНИ ПІНЧУК І МОЗ УКРАЇНИ ПРОПАГУЮТЬ "СЕКСУАЛЬНІ УРОКИ"
ЕКЗИСТЕНЦІЙНО-ПСИХОЛОГІЧНІ ОСНОВИ ПОРУШЕННЯ СТАТЕВОЇ ІДЕНТИЧНОСТІ ПІДЛІТКІВ
Батьківський, громадянський рух в Україні закликає МОН зупинити тотальну сексуалізацію дітей і підлітків
Відкрите звернення Міністру освіти й науки України - Гриневич Лілії Михайлівні
Представництво українського жіноцтва в ООН: низький рівень культури спілкування в соціальних мережах
Гендерна антидискримінаційна експертиза може зробити нас моральними рабами
ЛІВИЙ МАРКСИЗМ У НОВИХ ПІДРУЧНИКАХ ДЛЯ ШКОЛЯРІВ
ВІДКРИТА ЗАЯВА на підтримку позиції Ганни Турчинової та права кожної людини на свободу думки, світогляду та вираження поглядів
- Гідрологія і Гідрометрія
- Господарське право
- Економіка будівництва
- Економіка природокористування
- Економічна теорія
- Земельне право
- Історія України
- Кримінально виконавче право
- Медична радіологія
- Методи аналізу
- Міжнародне приватне право
- Міжнародний маркетинг
- Основи екології
- Предмет Політологія
- Соціальне страхування
- Технічні засоби організації дорожнього руху
- Товарознавство продовольчих товарів
Тлумачний словник
Авто
Автоматизація
Архітектура
Астрономія
Аудит
Біологія
Будівництво
Бухгалтерія
Винахідництво
Виробництво
Військова справа
Генетика
Географія
Геологія
Господарство
Держава
Дім
Екологія
Економетрика
Економіка
Електроніка
Журналістика та ЗМІ
Зв'язок
Іноземні мови
Інформатика
Історія
Комп'ютери
Креслення
Кулінарія
Культура
Лексикологія
Література
Логіка
Маркетинг
Математика
Машинобудування
Медицина
Менеджмент
Метали і Зварювання
Механіка
Мистецтво
Музика
Населення
Освіта
Охорона безпеки життя
Охорона Праці
Педагогіка
Політика
Право
Програмування
Промисловість
Психологія
Радіо
Регилия
Соціологія
Спорт
Стандартизація
Технології
Торгівля
Туризм
Фізика
Фізіологія
Філософія
Фінанси
Хімія
Юриспунденкция
Механізм дії ферментів
Після того, як була встановлена хімічна природа ферментів, підтвердилися припущення англійського хіміка А. Бруна, висловлене ще на початку XX ст., а згодом і вчених В. Анрі, Л. Міхаеліса та М. Ментен про те, що в основі ферментативного каталізу лежить утворення нестійкого проміжного фермент-субстратного комплексу, який згодом розпадається з утворенням продуктів реакції та вивільненням фермента в незміненому вигляді. Ці твердження лягли в основу теорії «жорсткої матриці» Е.Фішера про те, що активний центр фермента комплементарний субстрату, тобто підходить до нього як «ключ до замка». Згодом ця теорія була вдосконалена та доповнена в «гіпотезі індукованої відповідності» Д. Кошлендом, згідно з якою субстрат, взаємодіючи з активним центром фермента, викликає зміну його конформації, що призводить до формування фермент-субстратного комплексу (ЕS), сприятливого для хімічної модифікації субстрату; при цьому молекула субстрату теж змінює свою конформацію, що забезпечує високу ефективність ферментативної реакції.
Отже, базуючись на твердженнях Л. Міхаеліса та М. Ментен, процес ферментативного каталізу умовно можна розділити на три етапи: приєднання молекули субстрату (S) до фермента (Е); утворення проміжного фермент-субстратного комплексу та послідовне його перетворення на один або кілька перехідних (ЕS, ЕSх, ЕSхх) з подальшим утворенням нестабільного комплексу фермент-продукт (ЕР); вивільнення продуктів реакції (Р) та фермента (Е). Ці етапи можна описати наступним рівнянням і відобразити схематично (рис. 1.4):


Рис. 1.4. стадії ферментативного каталізу: Е – фермент, S – субстрат, ЕS – фермент-субстратний комплекс, Р – продукти реакції
Перша стадія – зближення та орієнтація субстрату відносно активного центра фермента з метою утворення фермент-субстратного комплексу – відбувається внаслідок зв’язування субстрату з активним центром ферменту водневими, електростатичними та гідрофобними взаємодіями, а в низці випадків – ковалентними та координаційними зв’язками. На цьому етапі має велике значення просторова конфігурація білкової молекули фермента, в якій жорсткі ділянки чергуються з еластичними лінійними відрізками, що надає молекулі фермента можливість динамічно змінюватися: приєднання субстрату до фермента змінює структуру активного центра останнього, його функціональні групи розміщуються так, що реакція може відбуватися. Утворення фемент-субстратного комплексу відбувається дуже швидко, тривалість цієї стадії залежить від концентрації субстрату, швидкості його дифузії до активного центра фермента. енергія активації при цьому змінюється несуттєво.
Друга стадія – перетворення первинного фермент-субстратного комплексу на один або кілька проміжних – характеризується послабленням зв'язків у субстраті, їх розривом або утворенням нових у результаті дії каталітичних груп фермента; формується молекула продукта. За рахунок утворення перехідних комплексів знижується енергія активації субстрату, що зумовлює зростання швидкості його перетворення. Ця стадія відбувається найповільніше та лімітує швидкість усього каталізу.
Третя стадія – відокремлення від комплексу продуктів реакції, які утворилися в процесі ферментативної реакції та перехід фермента у початковий стан. Тривалість цієї стадії наближається до першої та визначається швидкістю дифузії продукту в середовище.
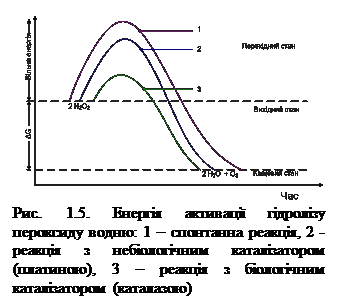 Як уже зазначалося, ферменти прискорюють біохімічні реакції за рахунок зниження енергії активації. Енергія активації – це енергія, необхідна для переходу молекул 1 моля речовини в активний (перехідний) стан при певній температурі. Іншими словами, це енергія, необхідна для запуску хімічної реакції. Фермент знижує енергію активації шляхом збільшення числа активованих молекул, які стають більш реакційно здатними на нижчому енергетичному рівні. Наприклад, при розщепленні пероксиду водню на воду та кисень енергія активації для досягнення перехідного (активного) стану дорівнює 18 ккал/моль. За участі платини енергія активації знижується до 12 ккал/моль, а під впливом фермента каталази – до 5 ккал/моль.
Як уже зазначалося, ферменти прискорюють біохімічні реакції за рахунок зниження енергії активації. Енергія активації – це енергія, необхідна для переходу молекул 1 моля речовини в активний (перехідний) стан при певній температурі. Іншими словами, це енергія, необхідна для запуску хімічної реакції. Фермент знижує енергію активації шляхом збільшення числа активованих молекул, які стають більш реакційно здатними на нижчому енергетичному рівні. Наприклад, при розщепленні пероксиду водню на воду та кисень енергія активації для досягнення перехідного (активного) стану дорівнює 18 ккал/моль. За участі платини енергія активації знижується до 12 ккал/моль, а під впливом фермента каталази – до 5 ккал/моль.
З рисунка 1.5 видно, що для досягнення збудженого стану вихідних речовин необхідно затратити найбільше енергії активації (рис. 1.5, 1); менше енергії вимагає реакція за участі небіологічного каталізатора (рис. 1.5, 2); і найменше енергії активації затрачається на реакцію за участі біологічного каталізатора (рис. 1.5, 3). Слід відмітити, що як каталізована ферментом, так і не каталізована ним реакція не залежно від її шляху має однакову величину стандартної зміни вільної енергії (ΔG).
Активний центр на всіх етапах ферментативного каталізу відіграє роль комплексного молекулярного механізму, який використовує різні хімічні перетворення, які сприяють перетворенню субстрату на продукт. на молекулярному рівні властивість контактних ділянок активного центра фермента специфічно зв'язувати субстрат і забезпечувати в такий спосіб їх взаємну орієнтацію та зближення так, щоб це було вигідно для дії каталітичних груп, називають ефектом зближення та орієнтації реагентів. Таке впорядковане розташування знижує енергію активації, що, своєю чергою, визначає каталітичну ефективність ферментів. Активний центр фермента також сприяє дестабілізації міжатомних зв’язків у молекулі субстрату, що полегшує перебіг хімічної реакції та утворення продуктів. Цю властивість активного центра називають ефектом деформації субстрату. Після зв'язування з активним центром молекула субстрату ніби розтягується. Місце деформації легше атакується, наприклад, молекулами води. Чим більша довжина міжатомного зв'язку в молекулі субстрату, тим менша енергія його розриву (знижується енгергія активації).
У залежності від ролі функціональних груп активного центра фермента розрізняють кислотно-основний і ковалентний каталіз.
Кислотно-основний каталіз характеризується участю в ферментативній реакції кислотних і/або основних груп. Так, радикали таких амінокислот як цис, тир, сер, ліз, глу, асп і особливо гіс можуть виступати як у ролі донорів протонів, так і їх акцепторів, надаючи ферментам властивостей універсальних каталізаторів, на відміну від небілкових каталізаторів, які проявляють або лише кислі, або лужні властивості. Прикладом такого каталізу може бути фермент алкогольдегідрогеназа печінки, яка містить іон Zn2+ та НАД+ у якості кофермента і каталізує реакцію окиснення етилового спирту: позитивно заряджений атом цинку сприяє від’єднанню протона від спиртової групи етанолу з утворенням негативно зарядженого атома кисню; негативний заряд перерозподіляється між атомом кисню та сусіднім атомом гідрогену, який потім у вигляді гідрит-іона переноситься на четвертий вуглецевий атом нікотинаміду з утворенням відновленої форми НАДН і оцтового альдегіду.
Ковалентний каталіз базується на утворенні ковалентних зв’язків між нуклеофільними (негативно зарядженими) або електрофільними (позитивно зарядженими) групами активного центра фермента та субстратом. Прикладом цього може слугувати дія серинових протеїназ (трипсину, хімотрипсину, тромбіну), до складу активного центра яких входить гідроксильна група серину та імідазольна група гістидину. Остання відтягує на себе протон від ОН-групи серину, внаслідок чого з’являється надлишкова електронна щільність на атомі кисню серину та полегшується нуклеофільна атака гідроксилом серину СО-групи субстрату. При цьому ацильний радикал від субстрату переноситься на сериновий залишок ферменту. Тоді атом гістидину здійснює нуклеофільну атаку на кисень ацильного похідного серину, внаслідок чого ацильна група відщеплюється і переноситься на молекулу води.
Максимальна активність ферменту спостерігається при оптимальних умовах перебігу реакції і зумовлена оптимальною конформацією молекули ферменту в цілому та активного центра зокрема. Тому навіть невеликі зміни умов, що впливають на зв'язування субстрату або конформацію третинної структури білка, будуть змінювати швидкість ферментативної реакції.
Читайте також:
- II. МЕХАНІЗМИ ФІЗІОЛОГІЧНОЇ ДІЇ НА ОРГАНІЗМ ЛЮДИНИ.
- V Процес інтеріоризації забезпечують механізми ідентифікації, відчуження та порівняння.
- Адвокатура — неодмінний складовий елемент механізму забезпечення прав людини.
- Аденогіпофіз, його гормони, механізм впливу
- Аденогіпофіз, його гормони, механізм впливу, прояви гіпер- та гіпофункцій.
- Адміністративно-командна система, її ознаки та механізм функціонування.
- Адміністративно-правові методи забезпечення економічного механізму управління охороною довкілля
- Аеробний механізм ресинтезу АТФ
- Акти застосування норм права в механізмі правового регулювання.
- Акціонерні товариства як механізм трансформації
- Альдостерон та механізми ренін-ангіотензину
- Аміноглікозиди (стрептоміцину сульфат, гентаміцину сульфат). Механізм і спектр протимікробної дії, застосування, побічні ефекти.
Переглядів: 9820
| <== попередня сторінка | | | наступна сторінка ==> |
| Номенклатура та класифікація ферментів | | | Кінетика ферментативних реакцій |
|
Не знайшли потрібну інформацію? Скористайтесь пошуком google: |
© studopedia.com.ua При використанні або копіюванні матеріалів пряме посилання на сайт обов'язкове. |