РЕЗОЛЮЦІЯ: Громадського обговорення навчальної програми статевого виховання
ЧОМУ ФОНД ОЛЕНИ ПІНЧУК І МОЗ УКРАЇНИ ПРОПАГУЮТЬ "СЕКСУАЛЬНІ УРОКИ"
ЕКЗИСТЕНЦІЙНО-ПСИХОЛОГІЧНІ ОСНОВИ ПОРУШЕННЯ СТАТЕВОЇ ІДЕНТИЧНОСТІ ПІДЛІТКІВ
Батьківський, громадянський рух в Україні закликає МОН зупинити тотальну сексуалізацію дітей і підлітків
Відкрите звернення Міністру освіти й науки України - Гриневич Лілії Михайлівні
Представництво українського жіноцтва в ООН: низький рівень культури спілкування в соціальних мережах
Гендерна антидискримінаційна експертиза може зробити нас моральними рабами
ЛІВИЙ МАРКСИЗМ У НОВИХ ПІДРУЧНИКАХ ДЛЯ ШКОЛЯРІВ
ВІДКРИТА ЗАЯВА на підтримку позиції Ганни Турчинової та права кожної людини на свободу думки, світогляду та вираження поглядів
- Гідрологія і Гідрометрія
- Господарське право
- Економіка будівництва
- Економіка природокористування
- Економічна теорія
- Земельне право
- Історія України
- Кримінально виконавче право
- Медична радіологія
- Методи аналізу
- Міжнародне приватне право
- Міжнародний маркетинг
- Основи екології
- Предмет Політологія
- Соціальне страхування
- Технічні засоби організації дорожнього руху
- Товарознавство продовольчих товарів
Тлумачний словник
Авто
Автоматизація
Архітектура
Астрономія
Аудит
Біологія
Будівництво
Бухгалтерія
Винахідництво
Виробництво
Військова справа
Генетика
Географія
Геологія
Господарство
Держава
Дім
Екологія
Економетрика
Економіка
Електроніка
Журналістика та ЗМІ
Зв'язок
Іноземні мови
Інформатика
Історія
Комп'ютери
Креслення
Кулінарія
Культура
Лексикологія
Література
Логіка
Маркетинг
Математика
Машинобудування
Медицина
Менеджмент
Метали і Зварювання
Механіка
Мистецтво
Музика
Населення
Освіта
Охорона безпеки життя
Охорона Праці
Педагогіка
Політика
Право
Програмування
Промисловість
Психологія
Радіо
Регилия
Соціологія
Спорт
Стандартизація
Технології
Торгівля
Туризм
Фізика
Фізіологія
Філософія
Фінанси
Хімія
Юриспунденкция
Вища нервова діяльність
4.1 Структура та функції кори великих півкуль
головного мозку
Вища нервова діяльність вивчає нервові механізми роботи мозку, які визначають поведінку тварин. Кора великих півкуль головного мозку та найближчі до неї підкіркові утворення відіграють першочергову роль у цих процесах.
Великі півкулі головного мозку у фізіологічному відношенні – це найбільш молодий відділ центральної нервової системи. Вони прогресивно розвивались від нижчих форм тварин до вищих (рис. 7).
Філогенетично найбільш стару нюхову частину кори мозку називають древньою корою (архікортекс). До старої кори (палеокортекса) відносять поясну звивину, звивину гіпокампа і мигдалик. Вся інша частина кори представляє нову кору (неокортекс). Древня кора добре розвинута у нижчих форм, наприклад у сумчастих, тоді як нова кора найбільш розвинута у вищих ссавців – у приматів. У тварин, що займають нижчі ступені еволюції кора мозку гладка. У більшості ссавців, включаючи копитних, хижаків та приматів, у корі є звивини та борозни, які в багато разів збільшують її поверхню та ускладнюють будову.
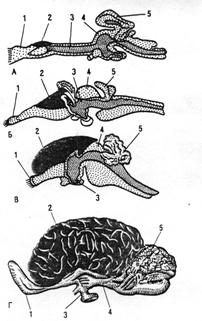
Рис. 7. Еволюція головного мозку хребетних:
А – головний мозок риби;
Б – головний мозок ящірки;
В – головний мозок кролика;
Г – головний мозок людини.
1 – нюхові долі;
2 – великі півкулі;
3 – проміжний мозок;
4 – середній мозок;
5 – мозочок.
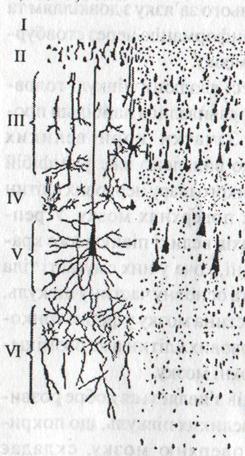 Рис. 8. Схема шестишарової
будови кори великих півкуль
Рис. 8. Схема шестишарової
будови кори великих півкуль
| Кора великих півкуль складається із нервових клітин, їх відростків і клітин нейроглії. Вона покриває всю поверхню великих півкуль. Товщина її 1,5 – 3 мм. Нервові клітини розташовані шарами. У вищих хордових тварин в найбільш розвинутих ділянках кори мозку морфологічно виділяють шість шарів (рис. 8). 1 шар – молекулярний, бідний нервовими клітинами, складається переважно із сплетінь нервових відростків. 2 шар – зовнішній зернистий, складається із дрібних, слабо диференційованих нервових клітин діаметром 4 – 8 мкм, кулястої, трикутної та багатокутної форми. 3 шар – має більшу товщину, ніж перші два шари, вміщує пірамідальні клітини різного розміру. 4 шар – внутрішній зернистий, складається із таких же клітин, як і другий шар. 5 шар – побудований з великих пірамідальних клітин. 6 шар – поліморфний, у ньому є різні за формою нейрони, такі як пірамідальні клітини, відростки яких доходять до 1,4 і 5 шарів. |
Функціонально нервові клітини кори мозку розподіляються на сенсорні, моторні та асоціативні. А нервові волокна кори мозку, які забезпечують взаємозв’язок між нейронами у межах однієї півкулі, називають міжцентральними, а ті, що з’єднують нейрони однієї півкулі з іншою – комісуральними, а ті, що проводять нервові імпульси у інші структури головного та спинного мозку – провідними. У корі великих півкуль виражена спеціалізація нервових центрів. Так, у кожній півкулі знаходиться моторна зона, у якій зосереджені рухові центри, що надсилають сигнали до окремих скелетних м’язів протилежної половини тіла. У цій же зоні знаходиться кінцева станція чутливих імпульсів, що надходять від рецепторів, які знаходяться у товщі м’язів, у сухожилках та суглобах. При цьому розрізняють видові особливості розміру та локалізації рухових центрів. Найбільш велика моторна зона у приматів, а найменша – у копитних, яким здебільшого властиві тільки прості рухи кінцівок. Двостороннє видалення кори мозку у вівці не викликало глибоких порушень рухових функцій одразу ж після операції; у м’ясоїдних же невдовзі після операції наступають порушення рухів, а у мавп – виникає параліч, який потім може пройти, але повного відновлення рухових функцій вже ніколи не відбудеться.
У м’ясоїдних моторна зона лежить навколо і в глибині крижової борозни, у овець і кіз займає область верхньої лобної звивини; у свиней розташована між вінцевою борозною та передньою гілкою сільвієвої борозни; у коней знаходиться збоку від латеральної клестовидної борозни, а також в області середньої гілки верхньої сільвієвої борозни.
У корі мозку є сенсорні зони: зорові, слухові, тактильні та інші. Зорова зона розташована в потиличних частках великих півкуль, слухова – у вилкових, нюхова – в амоновому розі древньої кори. В області задньої центральної звивини лежить тактильна зона, куди надходять імпульси від рецепторів шкіри, які реагують на дотик і тиск. У премоторній області є інтерорецептивна зона, яка отримує аферентні імпульси від внутрішніх органів. Подразнення або видалення цієї зони призводить до зміни дихання, роботи серця, просвіту судин, до порушення секреторної та моторної діяльності шлунково – кишкового тракту тощо.
І.П. Павлов є засновником вчення про вищу нервову діяльність. У його лабораторії вивчення функцій кори мозку здійснювалося різними методами. Розроблений ним метод умовних рефлексів дозволив у хронічних умовах вивчати процеси, що протікають у корі мозку, створити вчення про типи вищої нервової діяльності, а також про динамічну локалізацію функцій у корі великих півкуль головного мозку.
Встановлено, що коркова частина аналізатора включає центральне ядро, де проходить глибокий аналіз та синтез сигналів, що надходять і менш спеціалізовані периферичні елементи, які можуть виконувати функції центрального ядра, хоча не можуть замінити його повністю. Таким чином, у корі великих півкуль вищих тварин та людини сполучається принцип високої спеціалізації нервових центрів з їх гнучкістю і пластичністю, здатністю до динамічної перебудови своїх функцій.
4.1.1. Методи вивчення функцій кори
Найбільш простий та доступний метод вивчення вищої нервової діяльності тварин у різному стані – метод спостереження за їхньою поведінкою. Однак, цей метод є суб’єктивним і його можна використовувати разом з іншими методами.
Метод видалення окремих ділянок кори дає можливість з’ясувати функції різних її зон і кори в цілому.
У результаті видалення певної ділянки кори випадають або порушуються зв’язані з нею функції. Так, у собаки після видалення потиличної частки кори змінюються зорові функції: він не впізнає господаря, байдуже дивиться на кішку, що пробігла. Якщо видаляють вискову частку, у собаки порушується слухова функція і т.інше.
Повне видалення кори головного мозку у різних видів тварин показало, що чим вищий еволюційний розвиток тварини, тим важче відображається ця операція на її поведінці.
Наприклад, голуби з видаленою корою мозку зберігають координацію рухів. Вони можуть літати, клювати зерно. Але вибирати зерно серед непридатних для їжі предметів такі птахи невзмозі. Собаки після видалення кори можуть ходити, розпізнавати світло від темряви, реагувати на звуки, але всі реакції, що були набуті на протязі індивідуального життя, втрачаються.
Метод подразнення кори мозку полягає у тому, що під наркозом оголюють ділянку кори великих півкуль та на певні її місця наносять подразнення електричним струмом або хімічними речовинами. За допомогою цієї методики П. Фрітч та Є. Гітциг у 1870 році встановили локалізацію моторної зони у корі мозку собаки. Надалі методом подразнення було встановлено локалізацію рухової зони у корі мозку інших тварин. Метод подразнення при вивченні функції кори мозку використовують і в хронічних дослідах. Через отвори, просвердлені в окремих ділянках черепа, вводять електроди, що з’єднані з джерелом струму. Подразнення наносять після загоєння ран.
До сучасних методів вивчення функцій кори мозку відноситься електроенцефалографія або запис біострумів кори мозку. Існують багатоканальні електроенцефалографи, які дозволяють одночасно реєструвати потенціал дії з різних ділянок кори мозку.
Встановлено, що електроенцефалограми, які отримані з різних ділянок кори, відрізняються за частотою та амплітудою електричних потенціалів, а також залежать від функціонального стану нейронів кори великих півкуль головного мозку людини та тварин. Крива, що записана при відкритих очах та повній увазі до оточуючого, складається із швидких відхилень малої амплітуди ( Бета – ритм). Більш рідкі коливання, дещо більшої амплітуди, реєструються у зручному положенні при закритих очах ( Альфа – ритм ). Зовсім повільні, великі хвилі записують під час сну ( Гама – хвилі ).
Метод умовних рефлексів, розроблений І.П. Павловим, відкрив шляхи до вивчення головного мозку як складної цілісної функціональної системи.
У лабораторії І.П. Павлова вивчені умови та механізми утворення умовних рефлексів: слиновидільного, рухово–харчового і рухово–захисного. За допомогою умовних рефлексів можна суворо науково і об’єктивно аналізувати складну діяльність кори.
Функції кори мозку можна вивчати методом моделювання. Кібернетичні методи широко використовуються при аналізі роботи автоматів та машин. Можна моделювати і нервові процеси, які відбуваються у корі мозку, живого організму. Вони відкривають нові перспективи у вивченні функцій головного мозку та інших відділів центральної нервової системи. Аналізуючи результати, отримані при вивченні функцій кори півкуль головного мозку важливе місце займає теоретичний абстрактний розгляд проблем вищої нервової діяльності на основі апарату математичної логіки. Цей напрямок отримав назву теоретичної неврології.
4.1.2. Роль І.М. Сеченова та І.П. Павлова
у вивченні фізіології великих півкуль
Вивчення функцій кори мозку тривалий час було однією із найскладніших та нерозв’язаних проблем. Тільки на початку ХІХ століття вивчення функцій вищого відділу головного мозку було поставлено на наукову основу завдяки працям І.М. Сеченова та І.П.Павлова.
І.П.Павлов, базуючись на роботі І.М.Сеченова “Рефлекси головного мозку“, розробив вчення про умовні рефлекси. Високо оцінюючи роботу І.М.Сєченова як “геніальний зліт сеченовської думки ”, Павлов разом із своїми співробітниками експериментально довів, що і вищі відділи головного мозку функціонують по типу рефлексу. За допомогою методу умовних рефлексів І.П.Павлов створив вчення про вищу нервову діяльність. В основу цього вчення покладені три матеріалістичні принципи: детермінізму, аналізу і синтезу та структурності.
Принцип детермінізму полягає в тому, що психічна діяльність, як і всі явища природи, причинно обумовлена. Будь – який нервовий акт виникає не спонтанно, а під впливом того чи іншого подразника.
Принцип аналізу та синтезу полягає у наступному. У звичайних умовах на організм діє величезна кількість подразників. Центральна нервова система, головним чином кора головного мозку, здатна розкладати, аналізувати подразнення, що діють на рецептори, виділяти з них окремі елементи. Аналіз виявляється у здібності розрізняти форму різних предметів, їхні кольори, запахи, різницю температур та ін. Але за аналізом завжди іде синтез, з’єднання окремих елементів у певні комплекси. Відчуття від різних властивостей предмета зливаються у цілісний образ цього предмета. Наприклад, запах їжі, її форма, колір, смак синтезуються корою і виникає цілісне сприйняття певної їжі. Розвиток аналізуючої та синтезуючої здібності кори пов'язаний з пристосуванням організму до мінливих умов зовнішнього середовища.
Принцип структурності, пов'язаний з тим, що кожний нервовий процес виникає у певних морфологічних утвореннях. Такою структурою є нейрони мозку. Функція та структура взаємопов’язані, взаємно визначають одне одного. Сприймаючі області кори, де закінчуються аферентні шляхи від різних рецепторів, мають не тільки різну локалізацію, але й будову.
4.2. Умовні рефлекси
І.П. Павлов розробив методику вироблення умовних рефлексів, яка дозволила в умовах хронічного експерименту вивчити найскладнішу функцію кори півкуль головного мозку. Його класичні експерименти по умовно – рефлекторному виділенню травних соків дозволили відкрити, так звану “психічну фазу секреції”, яка подібно вродженим рефлексам, має рефлекторну природу та протікає за обов’язкової участі кори півкуль головного мозку та підкоркових утворень. Всі умовні рефлекси утворюються насамперед на базі вроджених, безумовних рефлексів. Разом з тим умовні рефлекси мають цілий ряд відмінностей від безумовних рефлексів.
4.2.1 Відмінності умовних рефлексів
від безумовних
1. Усі безумовні рефлекси є вродженими і передаються за спадковістю. Так, тільки що вилуплене з яйця курча одразу ж починає дзьобати, новонароджене теля – смоктати і т.д. Це безумовний харчовий рефлекс.
2. Умовні рефлекси виробляються при певних умовах у процесі всього життя тварин. Так, якщо цуценят до певного віку годувати тільки одним молоком, то в них виробляється стійкий умовний рефлекс на молоко: за наявності запаху молока у них виділяється слина та виникає рухова харчова реакція. При цьому хліб і м'ясо не викликають ніякої реакції. Але якщо 1–2 рази нагодувати цуценят м’ясом, то вигляд і запах м’яса викликатимуть слиновиділення.
3. Безумовні рефлекси – видові рефлекси. Всі кішки у разі небезпеки якої вони не можуть уникнути, вигинають спину та пирхають. Собаки при нападі на них гарчать та гавкають. Їжаки згортаються у клубок. Це захисні безумовні рефлекси. У різних видів тварин вони виявляються по різному, але у тварин одного виду безумовні рефлекси однакові.
4. Умовні рефлекси індивідуальні. Кожний організм по своєму реагує на один і той же подразник. Наприклад, одна тварина відгукується на одну кличку, а друга – на іншу. Для кожної особі існує свій комплекс умовних рефлексів, свій життєвий досвід.
5. безумовні рефлекси можуть здійснюватися і при відсутності вищих відділів центральної нервової системи. Якщо у кішки або собаки видалити кору великих півкуль, а потім вливати до рота кислоту, то і в цьому випадку у них буде виділятися слина. Усі безумовні рефлекси після видалення кори зберігаються. Отже, вони являють собою результат дії нижчих відділів центральної нервової системи. Умовні ж рефлекси зникають після видалення кори. Собаки без кори великих півкуль не реагують на кличку, не впізнають господаря, не виявляють рухових та слиновидільних реакцій на вигляд і запах їжі. Умовні рефлекси протікають за обов’язкової участі вищих відділів центральної нервової системи – кори великих півкуль та підкіркових структур головного мозку.
6. безумовні рефлекси виникають при подразненні певних рецепторних полів. Для кожного безумовного рефлекса є певна рецепторна зона і рефлекторна дуга. Умовні рефлекси можуть мати безліч рецепторних зон і рефлекторна дуга умовних рефлексів, її аферентна частина також різноманітна. Один і той же умовний рефлекс, наприклад слиновиділення, можна виробити при подразненні різних рецепторів: на світло лампочки, на звук дзвіночка, на дотик до шкіри тощо.
7. рефлекторна дуга безумовних рефлексів є постійною, а умовних рефлексів – тимчасовою. Не постійною є коркова частина рефлекторної дуги умовних рефлексів між нервовими центрами умовного та безумовного подразників.
4.2.2. Методики утворення умовних рефлексів
У лабораторії І.П. Павлова розроблені наступні методики відпрацювання умовних рефлексів: слиновидільна, рухово–захисна і рухово–харчова.
1. Суть слиновидільної методики полягає у тому, що у тварин попередньо хірургічним методом виводиться фістула протоку слинної залози, а потім вивчається слиновиділення при дії різних умовних та безумовних подразників. Так, вивчено характер слиновиділення у собаки при дії світлових та звукових подразників, які підкріплювались у процесі відпрацювання умовних рефлексів безумовними подразниками (м’ясом, хлібом і т.д.). Дана методика не може бути використана у широких масштабах для вивчення функцій кори мозку у сільськогосподарських тварин, оскільки потребує хірургічного втручання. Тому слиновидільна методика використовується насамперед на собаках в умовах експерименту (рис. 9).

Рис. 9. Схема постановки дослідів з вивчення умовних рефлексів:
1 – дзвоник; 2 – станок для фіксування тварини; 3 – годівниця;
4 – стіна; 5 – лампочка; 6 – слиновивід; 7 – система керування
2. Для вивчення вищої нервової діяльності сільськогосподарських тварин використовують рухово-захисну методику. Суть даної методики зводиться до того, що виробляють захисну реакцію на дію умовного подразника (світло, звук), який підкріплюється електричним подразником. З цією метою на путовий суглоб передньої кінцівки накладають електроди і наносять подразнення електричним струмом. Кожне больове подразнення супроводжується захисною реакцією у формі згинання кінцівки. В якості умовних подразників можна використовувати різні звукові, зорові, шкіряні подразники.
3. Для вивчення функцій кори мозку найрізноманітніших тварин можна використовувати рухово-харчову методику. Суть цієї методики зводиться до того, що при дії умовного подразника тварина рухається у пошуках харчового подразника. Тварина знаходиться у приміщенні, де вона може вільно рухатися. Безумовним подразником служить порція корму, яку кладуть у годівницю. Багаторазове сполучення того чи іншого умовного подразника з безумовним призводить до того, що тільки на звук або світло тварина йде до годівниці.
4.2.3. Механізм утворення умовних рефлексів
Рефлекси мають назву умовні тому, що виникають (і в природі, і в лабораторії) лише при певних умовах. Основними із них є:
1. Неодноразове сполучення дії умовного подразника з дією підкріплюючого безумовного подразника.
2. Дія умовного подразника повинна бути випереджаючою у часі. Тобто спочатку звучить дзвоник, а потім дається їжа. Причому протягом певного відрізку часу обидва подразники повинні діяти разом.
3. Відсутність сторонніх подразників. Тварина повинна знаходитися в ізольованій камері та реагувати лише на дію умовного і безумовного подразників.
4. Бадьорий стан організму тварини з високою збудливістю харчового центру безумовного подразнення. З цією метою тварину витримують на голодній дієті, а їжа повинна бути смачною. При цьому умовний подразник повинен бути оптимальної сили, а безумовний смачним, здатним викликати слиновиділення. Тварину, яку планують використовувати у досліді повинна бути молодою, здоровою, спокійною, вигуляною.
Так, при відпрацюванні слиновидільного рефлексу в ізольованій камері, де знаходиться піддослідна тварина запалюється лампочка і через 5-10 секунд подається їжа, тобто безумовний подразник. Причому протягом певного періоду збігається у часі дія обох подразників. Таке сполучення умовного сигналу та безумовного подразника повторюють декілька разів (за 1 дослід 8-10 сполучень). Через декілька сеансів вмикання лампочки буде викликати виділення слини без харчового підкріплення, що свідчить про відпрацювання слиновидільного умовного рефлексу.
Механізм утворення умовного рефлексу полягає у замиканні тимчасового нервового зв'язку у корі великих півкуль, між двома збудженими нервовими центрами: центром умовного та безумовного подразників.
При дії умовного подразника (світла) збуджуються рецептори сітківки ока і нервові імпульси по аферентним шляхам досягають зорового центру кори мозку. Майже одночасно із зоровим центром у корі мозку збуджується і харчовий центр кори під дією безумовного подразника - їжі, діючого на рецептори ротової порожнини. З харчового центру кори мозку нервові імпульси надходять у центр слиновиділення довгастого мозку і потім до слинних залоз, які виділяють слину. Таким чином, при відпрацюванні умовного слиновидільного рефлексу у корі мозку одночасно виникають два вогнища збудження: зорового та харчового центрів. При цьому центр безумовного подразника, тобто харчовий центр, збуджується у більшій мірі, ніж центр умовного подразника. Харчовому центру у корі мозку притаманна підвищена збудливість, сумація і по принципу домінанти притягування до себе збудження із центру умовного подразника. Таким чином, між центрами безумовного та умовного подразників утворюється тимчасовий зв'язок, який у подальшому забезпечує передачу нервових імпульсів між центрами умовного та безумовного подразників. Рефлекторна дуга умовно-рефлекторного слиновиділення буде складатися із наступних ланцюгів: рецептори сітківки, аферентні нервові волокна (зоровий нерв), зоровий центр кори мозку, тимчасовий зв'язок між зоровим та харчовим центрами у корі мозку, слиновидільний центр довгастого мозку, еферентні нервові волокна (блукаючий нерв) та слинні залози. Слід відмітити, що рефлекторні дуги умовного та безумовного рефлексів слиновиділення мають різні рецепторні початки, а закінчуються загальним центробіжним шляхом. Так, рефлекторна дуга безумовного рефлексу слиновиділення починається з рецепторів ротової порожнини, а умовного рефлексу слиновиділення, на увімкнення лампочки починається з рецепторів сітківки очей. У першому випадку збудження досягає харчового центру кори по аферентним нервовим волокнам через центр слиновиділення довгастого мозку. А під дією світла збудження досягає харчового центру кори через рецептори сітківки очей, зоровий центр кори і тимчасовий зв'язок між зоровим та харчовим центрами у корі мозку. Загальний центробіжний шлях слиновидільного рефлексу включає: харчовий центр кори мозку, слиновидільний центр довгастого мозку, еферентні нервові волокна та слинні залози.
Є й інші теорії утворення тимчасових зв'язків у корі мозку при відпрацюванні умовних рефлексів. П.К Анохін вважає, що при сполученні умовного та безумовного подразників трапляються деякі зміни у розташуванні азотистих основ молекул рибонуклеїнової кислоти і в наслідок зміни коду РНК в аксоплазмі утворюються нові своєрідні білкові молекули, особливо чутливі до певних нервових імпульсів. Ці молекули є "зберігачами" зв'язку, що утворився між центрами умовного та безумовного подразників.
В утворенні стійкого стану постійної провідності у тимчасових зв'язках, що утворилися, велике значення має циркуляція імпульсів по кільцевих системах кори. Наприклад, перехідні клітини п'ятого слою від свого низхідного аксону віддають бокові гілочки, які підіймаються у верхні шари кори і закінчуються у вставних нейронах другого та третього шарів. По цих шляхах імпульси від пірамідної клітини можуть частково повертатися до вставних нейронів і через них повторно збуджувати пірамідну клітину. Можливо, вихід імпульсів у ці бокові замкнуті кола відбувається при достатній інтенсивності збудження, або при підвищеній збудливості нервових елементів даного шляху. Це трапляється при сполученні умовного подразника з безумовним, коли зливаються два потоки імпульсів. Встановлена безперервна циркуляція імпульсів збудження по такій замкнутій кільцевій системі буде, у свою чергу, підтримувати стан підвищеної збудливості і тих нейронів, у синапсах яких замикається тимчасовий зв'язок.
Читайте також:
- VІ. Практична діяльність учнів.
- Адвокатська діяльність
- Актове діловодство. Діяльність судів. Ведення актових книг
- Англійська система фізичного виховання. Діяльність Томаса Арнольда.
- Антимонопольна діяльність держави
- Антимонопольна діяльність держави
- Антимонопольна діяльність держави.
- Аудиторська фірма — юридична особа, створена відповідно до законодавства, яка здійснює виключно аудиторську діяльність.
- Банківська діяльність
- Банківська діяльність у сфері надання фінансових послуг
- Банківська діяльність у сфері надання фінансових послуг.
- Бізнес як діяльність.
| <== попередня сторінка | | | наступна сторінка ==> |
| Форми поведінки тварин | | | Концепція П.К. Анохіна (1964) про формування реакцій поведінки |
|
Не знайшли потрібну інформацію? Скористайтесь пошуком google: |
© studopedia.com.ua При використанні або копіюванні матеріалів пряме посилання на сайт обов'язкове. |