РЕЗОЛЮЦІЯ: Громадського обговорення навчальної програми статевого виховання
ЧОМУ ФОНД ОЛЕНИ ПІНЧУК І МОЗ УКРАЇНИ ПРОПАГУЮТЬ "СЕКСУАЛЬНІ УРОКИ"
ЕКЗИСТЕНЦІЙНО-ПСИХОЛОГІЧНІ ОСНОВИ ПОРУШЕННЯ СТАТЕВОЇ ІДЕНТИЧНОСТІ ПІДЛІТКІВ
Батьківський, громадянський рух в Україні закликає МОН зупинити тотальну сексуалізацію дітей і підлітків
Відкрите звернення Міністру освіти й науки України - Гриневич Лілії Михайлівні
Представництво українського жіноцтва в ООН: низький рівень культури спілкування в соціальних мережах
Гендерна антидискримінаційна експертиза може зробити нас моральними рабами
ЛІВИЙ МАРКСИЗМ У НОВИХ ПІДРУЧНИКАХ ДЛЯ ШКОЛЯРІВ
ВІДКРИТА ЗАЯВА на підтримку позиції Ганни Турчинової та права кожної людини на свободу думки, світогляду та вираження поглядів
- Гідрологія і Гідрометрія
- Господарське право
- Економіка будівництва
- Економіка природокористування
- Економічна теорія
- Земельне право
- Історія України
- Кримінально виконавче право
- Медична радіологія
- Методи аналізу
- Міжнародне приватне право
- Міжнародний маркетинг
- Основи екології
- Предмет Політологія
- Соціальне страхування
- Технічні засоби організації дорожнього руху
- Товарознавство продовольчих товарів
Тлумачний словник
Авто
Автоматизація
Архітектура
Астрономія
Аудит
Біологія
Будівництво
Бухгалтерія
Винахідництво
Виробництво
Військова справа
Генетика
Географія
Геологія
Господарство
Держава
Дім
Екологія
Економетрика
Економіка
Електроніка
Журналістика та ЗМІ
Зв'язок
Іноземні мови
Інформатика
Історія
Комп'ютери
Креслення
Кулінарія
Культура
Лексикологія
Література
Логіка
Маркетинг
Математика
Машинобудування
Медицина
Менеджмент
Метали і Зварювання
Механіка
Мистецтво
Музика
Населення
Освіта
Охорона безпеки життя
Охорона Праці
Педагогіка
Політика
Право
Програмування
Промисловість
Психологія
Радіо
Регилия
Соціологія
Спорт
Стандартизація
Технології
Торгівля
Туризм
Фізика
Фізіологія
Філософія
Фінанси
Хімія
Юриспунденкция
МЕМБРАНИ МІКРОБНИХ КЛІТИН. Цитоплазматична мембрана.
Фарбування за Грамом.Диференційоване фарбування бактерій генціанвіолетом було запропоновано у 1884 р. датськиі фармакологом Г.Х. Грамом. У мікробіології забарвлення за Грамом є важливою таксономічною ознакою, з якою корелюють інши властивості бактерій. Суть методу полягає в тому, що при фарбуванні бактерій генціанвіолетом (кристалвіолетом, метилвіо летом) фарба з йодом утворюють сполуку, що утримується клітинами при обробці їх спиртом. Такі бактерії забарвлені в синьофіолетовий колір і їх називають грампозитивними.
Будова і хімічний склад клітинних стінок прокаріот.
Хімічний склад клітинних стінок мікроорганізмів спочатку привертав увагу дослідників у галузі систематики. Основою тому були дані про якісні відмінності у складі клітинних стінок між еукаріотними та прокаріотними мікроорганізмами, а також серед прокаріотів — між грампозитивними та грамнегативними бактеріями.
Бактерії, які знебарвлюються при обробці спиртом, називаються грамнегативними. їх потім дофарбовують контрастною фарбою (фуксином).
У 1978 р. Н.Е.Гіббонс та Р.Г.Е.Муррей запропонували грамнегативні істинні бактерії (еубактерії) виділити у відді Грацилікутних (Gracilicutes), а грампозитивні — у відділ Фірмікутних (Firmicutes). Але терміни "фірмікутні", "грацилікутні" бактерії не мають широкого використання в мікробіології.
Пептидоглікан (глікопептид, мукопептид, муреїн).Основним компонентом клітинної стінки бактерій є пептидогліка (глікопептид, мукопептид, муреїн). Пептидоглікан виявлен тільки у прокаріот. Винятком є еубактерії, що не мають клітинної стінки (мікоплазми, L-форми), та архебактерії — деякі метанеутворювальні та галофіли (Halobacterium, Halococcus). Для гало-
фільних бактерій наявність міцної клітинної стінки не обов'язкова, оскільки вміст їх клітин є ізоосмотичним з навколишнім середовищем.
Специфічний гетерололімер пептидоглікан складається: із залишків N -ацетилглюкозаміну та N-ацетилмурамової кислоти, з'єднаних між собою в-1,4-глікозидними зв'язками. N-ацетилглюкозамін є похідною сполукою глюкози, в якій гідроксильна група при другому атомі вуглецю заміщена на аміногрупу. N-ацетилмурамова кислота — це ефір N-ацетилглюкозаміну та .D-молочної кислоти {рис. 2.3);
із діамінокислот, з яких найчастіше зустрічаються мезо-діамінопімелінова кислота, LL-діамінопімелінова кислота, лізин, орнітин. Наявність таких амінокислот з двома аміногрупами має принципове значення для просторової організації пептидоглікану. Вони забезпечують утворення двох пептидних зв'язків між пептидними угрупованнями в молекулі;

Рис.2.3.Основні складові гетерополімеру пептидоглікану
з інших амінокислот (D- та L-аланін, D-глутамінова кислота, L-серин, гліцин).
Фрагмент пептидоглікану показано на рис.2.4.. За допомогою пептидних місточків гетерополімерні ланцюги зв'язані між собою в мішкоподібну гігантську молекулу — муреїновий мішок (муреїнова сітка).
Муреїновий мішок виконує функцію опорного каркаса клітинної стінки. За будовою цього каркаса, а також за вмістом інших речовин у клітинній стінці грампозитивні бактерії відрізняються від грамнегативних.. Клітинна стінка грампозитивних бактерій.У грампозитивних бактерій частка муреїнової сітки становить 30-70 % сухої маси клітинної стінки (завтовшки 40 шарів). Замість мезо-діамінопімелінової кислоти часто міститься LL-діамінопімелінова кислота або лізин. У клітинній стінці грампозитивних бактерій полісахариди, якщо вони є, зв'язані між собою ковалентно. Вміст ліпідів і білків невисокий. У білках клітинних стінок грампозитивних бактерій набір амінокислот менший (4-12), ніж у грамнегативних (містяться практично всі амінокислоти, з яких складаються білки).

Рис. 2.4.Структура пептидоглікану Escherichia coli:
N-АцМур — N-ацетилмурамова кислота; N-АцГлю — N-ацетилглюкозамін; Ала — аланін; Глу — глутамінова кислота; m-Дпм — мезо-діамінопімелінова кислота; ▼ — лізоцим
Характерною особливістю грампозитивних бактерій є наявність у клітинній стінці тейхоєвих кислот. Тейхоєві кислоти — це ланцюги, які складаються з 8-50 залишків гліцерину чи рибітолу, зв'язаних між собою фосфатними місточками. У молекулі тейхоєвої кислоти поліол може містити моносахариди як замісники. Деякі з тейхоєвих кислот містять еритритол чи маніт. Припускається, що тейхоєві кислоти зв'язані з муреїном через фосфат за типом аміду. У складі тейхоєвих кислот деяких грампозитивних бактерій містяться жирні кислоти, які утворюють ефірні зв'язки з гліцериновими залишками. їх називають л і п о т е й х о є в и м и кислотами. Тейхоєві кислоти містяться в клітинах у значних кількостях. У деяких бактерій вони становлять більше половини маси клітинної стінки.
Функції тейхоєвих кислот:
фосфатні групи тейхоєвих кислот є місцем зв'язування катіонів магнію, необхідного для багатьох ензиматичних і фізико-хімічних процесів, що проходять на цитоплазматичній мембрані;
тейхоєві кислоти беруть участь у регуляції активності авто-літичних ферментів;
було показано, що цукрові компоненти тейхоєвих кислот є відповідальними за зв'язування фатів з клітинною стінкою. Якщо тейхоєва кислота з будь-яких причин втрачає глікозильні замісники, то бактеріальна клітина стає фагорезистентною (фагостійкою);
ліпотейхоєві кислоти беруть участь в імунологічних реакціях.
Клітинна стінка грамнегативних бактерій.У грамнегативних бактерій муреїнова сітка є одношаровою і становить менше 10 % сухої маси клітинної стінки. Муреїн містить тільки .мезо-діамінопімелінову кислоту і не містить лізину. У складі клітинних стінок грамнегативних бактерій тейхоєві кислоти не виявлено.
У всіх грамнегативних бактерій зверху одношарового чи найбільше двошарового муреїнового мішка розміщується зовнішній шар клітинної стінки. Це так звана зовнішня мембрана, що складається з білків, фосфоліпідів і ліпополісахаридів (ЛПС) (рис.2.5).
З муреїном, очевидно, ковалентно через діамінопімелінову кислоту зв'язані ліпопротеїни. Вони орієнтовані своїми ліпофільними кінцями назовні і таким чином закріплені в ліпофільному подвійному шарі (завдяки гідрофобній взаємодії). У цьому ж шарі містяться фосфоліпіди та гідрофобні кінці ліпополісахаридів. Гідрофільні кінці ЛПС орієнтовані назовні.
 Ліпополісахариди — складні молекули з молекулярною масою більш як 10 000. Вони складаються з трьох частин — ліпіду А, ядра {кор, серцевинна зона) таО-специфічного бокового ланцюга. ЛПС Salmonella typhimurium та інших ентеробактерій досліджені досить повно.
Ліпополісахариди — складні молекули з молекулярною масою більш як 10 000. Вони складаються з трьох частин — ліпіду А, ядра {кор, серцевинна зона) таО-специфічного бокового ланцюга. ЛПС Salmonella typhimurium та інших ентеробактерій досліджені досить повно.
Рис. 2.5. Модель будови клітинної стінки грамнегативних бактерій:
Праворуч — ліпополісахаридна молекула; Глю — глюкоза; Глю-N — глюко-
замін; NA=N-AцГлю — N-ацетилглкжозамін; Гал — галактоза; Геп — гепто-
за; КДО — 2-кето-З-дезоксиоктонова кислота; М — муреїн; ЗМ — зовнішня
мембрана; ПМ — плазматична мембрана;ПП — периплазматичний простір
Ліпід А складається з глюкозаміндисахариду, до гідроксильних груп якого ефірними зв'язками приєднані жирні кислоти (С12, С14, С16), Ця частина молекули має гідрофобні властивості. Далі міститься R-серцевинна зона— трисахарид, що складається з трьох залишків 2-кето-З-дезоксиоктонової кислоти (КДО) і який зв 'язаний також з фосфоетаноламіном. Далі йдуть дві молекули гептози і зовнішня серцевинна зона. Остання складається з розгалуженого ланцюга, що містить глюкозу, галактозу і N-ацетилглюкозамін. Ця базова структура є однаковою у всіх сальмонел. До серцевинної зони прилягає О-специфічний боковий ланцюг. Це довгі ланцюги, що складаються з повторюваних олігосахаридів, які можуть містити галактозу, манозу, рамнозу, абеквозу, фукозу та інші моносахариди у послідовності, що варіює від штаму до штаму.
ЛПС набули великого значення в бактеріологічній діагностиці і розпізнаванні епідемій. Виявилось, що збудники різних захворювань відрізняються один від одного О-специфічними боковими ланцюгами. Незначні відмінності в їх складі можуть бути виявлені за допомогою імунологічних методів. За серологічними реакціями у роді Salmonella вдалося виділити понад тисячу видів і штамів. Є так звані місцеві раси сальмонел, які можна ідентифікувати за імунохімічними особливостями. Це часто дає можливість встановити, де відбулося зараження хворого чи звідки почала поширюватися епідемія. Наприклад, можна встановити, де хворий отримав інфекцію — у південно-американському чи східно-азіатському регіоні.
Функції зовнішньої мембрани. Зовнішня мембрана грамнегативних бактерій виконує не тільки механічні, а й важливі фізіологічні функції. В її подвійний ліпідний шар, що складається з ліпіду А, полісахаридів і фосфоліпідів вбудовані білки, які пронизують цей шар наскрізь. Ці трансмембранні білки називаються п о р и н а м и. Порини пропускають через мембрану гідрофільні низькомолекулярні речовини (до молекулярної маси близько 6 000).
Зовнішня мембрана прилягає до муреїнового шару і зв'язана з ним ліпопротеїнами. Очевидно, муреїновий шар є проникним для різних сполук. Проміжок між муреїном і плазматичною мембраною називають л ер ця л а з мати ч ним простором. У ньому містяться ферменти, в тому числі і деполімерази (протеїнази, нуклеази), периферійні білки і так звані зв'язувальні білки. Останні беруть участь у перенесенні деяких субстратів у цитоплазму і є рецепторами хемотаксичних сигналів. Периплазматичний простір, очевидно, відіграє певну роль в осморегуляції.
Дія лізоциму та пеніциліну.Структура клітинної стінки та муреїну була встановлена у зв'язку з вивченням дії лізоциму та пеніциліну на бактерії. Відкритий англійським мікробіологом А. Флемінгом у 1922 р. лізоцим є бактерицидним ферментом, що міститься в яєчному білку, носовому слизі, в сльозовій рідині. Лізоцим виділили також з бактерій {Е. coli, Streptomyces) і бактеріофагів. При дії лізоциму на суспензію грампозитивних бактерій спостерігали швидке її просвітлення. Так, Mіcrococcus luteus лізується (розчиняється) вже у концентрації 1 мкг лізоциму на 1 мл. Для лізису клітин Bacillus megaterlum необхідною є концентрація 50 мкг/мл, а грамнегативні бактерії розчиняються тільки за наявності в суспензії ЕДТА.
Лізоцим розриває в муреїні глікозидний зв'язок між першим вуглецевим атомом N-ацетилмурамової кислоти і четвертим вуглецевим атомом N-ацетилглюкозаміну . При цьому полісахаридні ланцюги розщеплюються до дисахаридних фрагментів. Отже, лізоцим є N-ацетилмурамідазою.
Слід зазначити, що повному руйнуванню бактеріальних клітин можна запобігти, здійснюючи лізис в ізотонічному чи слабкогіпертонічному розчині (0,1-0,2 М сахарози). У цих умовах під дією лізоциму з клітин утворюються надзвичайно чутливі до осмотичних умов округлі протопласти. У гіпертонічних та ізотонічних розчинах протопласти стабільні, у гіпотонічних — лопаються. Протопластами слід називати тільки такі округлі клітини, в яких немає ніяких залишків клітинної стінки, тобто не можна виявити ні мурамової кислоти, ні специфічної для клітинної стінки діамінопімелінової кислоти. Лізис клітинної стінки не супроводжується порушеннями метаболізму.
Крім лізоциму, є ряд інших ферментів, що руйнують муреїновий каркас, наприклад, муроендопептидази.
Антибіотик пеніцилін діє переважно на грампозитивні бактерії (пневмококи та стафілококи), а також на деякі грамнегативні (гонококи, менінгококи, ентеробактерії), вбиваючи їх. Але бактерицидній дії піддаються тільки клітини, які ростуть. Клітини, що перебувають у стані спокою, залишаються живими. Найцікавіший феномен, який спостерігається під дією пеніциліну, — це поява так званих L - ф о р м, які утворюються з нормальних бактеріальних клітин у результаті незбалансованого росту в довжину та ширину. При цьому вихідні палички збільшуються в об'ємі в багато разів. Якщо діяти пеніциліном на клітини, що ростуть, у гіпотонічному розчині вони лопаються, В ізо- та гіпертонічних розчинах палички перетворюються на шароподібні утворення, які називаються L-формами або сферопластами. Від протопластів вони відрізняються тим, що зберігають залишки клітинної стінки. Пеніцилін порушує процес утворення клітинної стінки.
Клітина будь-якого організму містить різні мембрани, що розрізняються морфологічно та функціонально. Безпосередньо під клітинною стінкою розміщена цитоплазматична мембрана {плазматична мембрана, плазмалема).
Ця мембрана покриває цитоплазму. У житті клітини вона має велике значення, виконуючи роль не тільки структурного морфологічного компонента. Мембрана є осмотичним бар'єром організму, який регулює осмотичний тиск всередині клітини. Через мембрану здійснюється вибірковий транспорт поживних речовин із середовища в клітину та вихід із неї продуктів обміну — метаболітів. Мембрана є місцем, де відбувається синтез деяких клітинних структур, зокрема клітинної стінки та капсули. У мембрані локалізований і з мембраною асоційований ряд ферментів (ферменти перенесення електронів та окисного фосфорилювання, які в еукаріот містяться в мітохондріях, у бактерій локалізовані всередині чи на поверхні плазматичної мембрани; компоненти електрон-транспортного ланцюга — дихального ланцюга — містяться тільки в мембранах). У мембрані міститься весь фотосинтетичний апарат у пурпурових бактерій. Цілком можливо, що на мембрані локалізується центр реплікації ДНК.
Мембрани можна виділити, якщо піддати осмотичному тиску протопласти, одержані за допомогою лізоциму. Мембрана багата на ліпіди, особливо фосфоліпіди (табл. 2.1.). Маючи у складі всього 8-15 % сухої речовини клітини, мембрани містять 70-90 % усіх її ліпідів. За своєю будовою мембрани мікробних, рослинних і тваринних клітин дуже подібні. Це дає підставу говорити про універсальну "елементарну мембрану".
Класичною, однією з перших моделей будови мембран, була модель Даніеллі—Даусона—Робертсона. Мембрана складається з подвійного шару ліпідів, що міститься між тонкими шарами білка. Внутрішній шар мембрани складається з ліпідів, у яких, як відомо, є полярний кінець (гідрофільний), здатний до іонізації, та неполярний (гідрофобний), що за своєю хімічною природою являє собою вуглеводневий ланцюг. Ліпіди в мембрані орієнтовані вуглеводневими кінцями один до одного, полярними кінцями — назовні, тобто утворюють подвійний шар (бімоле-
кулярний шар). З полярними кінцями стикується мономолекулярний шар неліпідної природи. Здебільшого це білок.
Вперше припущення про те, що в мембрані присутні білки, було висловлено англійськими вченими Дж.Ф. Даніеллі та X. Даусоном у 1935 р. для пояснення низького поверхневого натягу клітинних мембран. Оскільки на межі поділу масло— вода повинен виникати великий поверхневий натяг, ці вчені прийшли до висновку, що гідрофобність ліпідних компонентів повинна компенсуватись якимсь гідрофільним білком. У 1959 р. на основі робіт американського вченого Дж. Д. Робертсона (за допомогою електронної мікроскопії було виявлено два електронощільних шари мембрани, розділених менш щільною ділянкою) була сформульована гіпотеза елементарної мембрани. Наявність електронощільних зовнішніх шарів мембрани
Таблиця 2.1. Склад мембран деяких бактерій
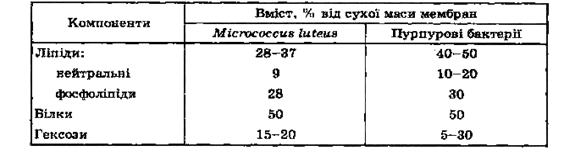
пояснювалась тим, що з гідрофільними поверхнями, що утворюються ліпідними молекулами, зв'язаний білок. Але проти цієї моделі свідчили результати електронно-мікроскопічних досліджень препаратів мембран, які були одержані методом заморожування-сколювання: виявилось, що білкові молекули розміщуються не тільки на поверхні мембрани, а є і такі, що пронизують її наскрізь. Рідинно мозаїчна модельструктури мембрани була запропонована у 1972 р.
Г. Ніколсоном та С.Д. Сінгером (рис. 2.6). Згідно з цією моделлю, що
отримала в наш час загальне визнання, білки можна уявити як айсберги, що плавають у ліпідному морі.
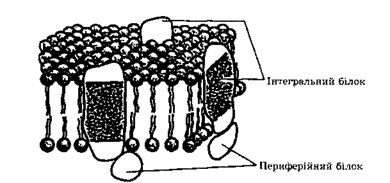
Рис. 2.6.Рідинно-мозаїчна модель плазматичної мембрани Ніколсона—Сінгера.
У подвійний шар ліпідів занурені інтегральні білки. Периферійні білки розміщені на поверхні мембрани
Мембранні білки.Існують два типи мембранних білків: Інтегральні та периферійні (див. рис. 2.6).
Периферійні білки легко вимиваються з мембрани м'якими детергентами або навіть дистильованою водою, вони зв'язані з поверхнею мембрани. На відміну від периферійних інтегральні білки пронизують товщу мембрани наскрізь. Як правило, інтегральні білки перебувають у комплексі з ліпідами. Білки цих двох типів повинні різнитись між собою за розміщенням гідрофобних амінокислотних залишків. Поверхня периферійних білків гідрофільна (ці білки розчинні у воді), а гідрофобні амінокислотні залишки занурені всередину білкової глобули. В інтегральних білках гідрофобні залишки локалізовані на поверхні, забезпечуючи максимальну взаємодію з неполярним середовищем всередині мембрани. Проте у деяких інтегральних білків полярні групи теж розміщені на поверхні і взаємодіють з полярними групами ліпідів і з периферійними білками.
За біологічними функціямимембранні білки умовно поділяються на три групи:
з притаманною ферментативною активністю;
специфічно зв'язують різні речовини, необхідні для клітини, тобто білки, яким притаманна рецепторна функція (білки-пермеази);
структурні, але вони хімічно мало досліджені. Відомо, що всі вони слабкорозчинні у воді із-за наявності великих гідрофобних ділянок. Це створює умови для утворення міцних структур з ліпідами — ліпопротеїдів.
Мембранні вуглеводи. Вільних вуглеводів у клітинних мембранах мало, більшість вуглеводних залишків входить до складу гліколіпідів і глікопротеїдів. В їх складі виявлені одні й ті самі моносахариди: галактоза, глюкоза, N-ацетилглюкозамін, N-ацетилгалактозамін, фукоза, маноза, ксилоза.
Ліпіди мембран. Ліпіди представлені в основному фосфоліпідами та гліколіпідами. З фосфоліпідів найчастіше у бактерій зустрічаються фосфатидилгліцерин і фосфатидилетаноламін. Фосфатидилхолін, фосфатидилінозит зустрічаються рідше. Фосфоліпіди містять у своїй молекулі фосфор, зв'язаний двома ефірними зв'язками. До складу фосфоліпідів входить один спільний компонент — гліцерин, з яким з'єднані ефірним зв 'язком дві жирні кислоти з довгим ланцюгом, а також фосфорвмісна сполука. Наявність двох неполярних залишків жирних кислот у складі мембранних структур є їх характерною особливістю. У бактеріальних культур ліпіди, у тому числі й фосфоліпіди, містять переважно насичені жирні кислоти (хоча є і ненасичені). Присутні в мембранах і жирні кислоти з розгалуженим ланцюгом (особливо у сарцин і мікрококів).
Вважається, що однією з ознак, які відрізняють бактеріальні ліпіди від ліпідів інших мікроорганізмів, є відсутність стеринів у бактерій.
Крім ліпідів, що містять гліцерин, у деяких сполуках присутній етиленгліколь. Такі ліпіди називають диольними. У великих концентраціях диольні ліпіди руйнують мембрани. Але в дуже обмежених кількостях вони лише змінюють їх властивості, наприклад, підвищують проникність для невеликих іонів і молекул. Очевидно, клітини використовують цю властивість. Так, у період швидкого росту вони інтенсивно синтезують диольні ліпіди. Коли ріст сповільнюється, синтез диольних ліпідів припиняється.
Гліколіпіди є вуглеводними похідними ліпідів.
У структуру мембран входять також іони двовалентних металів. Передбачається, що вони утворюють з фосфоліпідами хелатні комплекси і тим самим надають мембрані необхідної міцності за рахунок більшої компактності.
Читайте також:
- Б- не збуджена ділянка мембрани , на яку діють електричні струми збудженої ділянки. Стрілками показано напрям струмів, кружечками – дійсне переміщення іонів.
- Модельні ліпідні мембрани
- Рідинні мембрани.
- Тема: Вступ. Предмет, методи і завдання патоморфології та патофізіології. Ушкодження клітин.
- Тотіпотентність рослинних клітин. Тотіпотентність тваринних клітин раннього зародку.
- Трансплантація ядер і реконструювання клітин.
- Транспорт іонів та інших речовин через мембрани, його види, механізм реалізації.
- Цитоплазматична чоловіча стерильність.
- ЧЕРЕЗ БІОЛОГІЧНІ МЕМБРАНИ
| <== попередня сторінка | | | наступна сторінка ==> |
| Поверхневі структури клітинної стінки бактерій. | | | Мембранні утворення у прокаріот. |
|
Не знайшли потрібну інформацію? Скористайтесь пошуком google: |
© studopedia.com.ua При використанні або копіюванні матеріалів пряме посилання на сайт обов'язкове. |