РЕЗОЛЮЦІЯ: Громадського обговорення навчальної програми статевого виховання
ЧОМУ ФОНД ОЛЕНИ ПІНЧУК І МОЗ УКРАЇНИ ПРОПАГУЮТЬ "СЕКСУАЛЬНІ УРОКИ"
ЕКЗИСТЕНЦІЙНО-ПСИХОЛОГІЧНІ ОСНОВИ ПОРУШЕННЯ СТАТЕВОЇ ІДЕНТИЧНОСТІ ПІДЛІТКІВ
Батьківський, громадянський рух в Україні закликає МОН зупинити тотальну сексуалізацію дітей і підлітків
Відкрите звернення Міністру освіти й науки України - Гриневич Лілії Михайлівні
Представництво українського жіноцтва в ООН: низький рівень культури спілкування в соціальних мережах
Гендерна антидискримінаційна експертиза може зробити нас моральними рабами
ЛІВИЙ МАРКСИЗМ У НОВИХ ПІДРУЧНИКАХ ДЛЯ ШКОЛЯРІВ
ВІДКРИТА ЗАЯВА на підтримку позиції Ганни Турчинової та права кожної людини на свободу думки, світогляду та вираження поглядів
- Гідрологія і Гідрометрія
- Господарське право
- Економіка будівництва
- Економіка природокористування
- Економічна теорія
- Земельне право
- Історія України
- Кримінально виконавче право
- Медична радіологія
- Методи аналізу
- Міжнародне приватне право
- Міжнародний маркетинг
- Основи екології
- Предмет Політологія
- Соціальне страхування
- Технічні засоби організації дорожнього руху
- Товарознавство продовольчих товарів
Тлумачний словник
Авто
Автоматизація
Архітектура
Астрономія
Аудит
Біологія
Будівництво
Бухгалтерія
Винахідництво
Виробництво
Військова справа
Генетика
Географія
Геологія
Господарство
Держава
Дім
Екологія
Економетрика
Економіка
Електроніка
Журналістика та ЗМІ
Зв'язок
Іноземні мови
Інформатика
Історія
Комп'ютери
Креслення
Кулінарія
Культура
Лексикологія
Література
Логіка
Маркетинг
Математика
Машинобудування
Медицина
Менеджмент
Метали і Зварювання
Механіка
Мистецтво
Музика
Населення
Освіта
Охорона безпеки життя
Охорона Праці
Педагогіка
Політика
Право
Програмування
Промисловість
Психологія
Радіо
Регилия
Соціологія
Спорт
Стандартизація
Технології
Торгівля
Туризм
Фізика
Фізіологія
Філософія
Фінанси
Хімія
Юриспунденкция
Загальні відомості
Формені елементи крові.
|
 Еритроцити (erythrocytus), або червонокрівці (Рис.2.1), у ссавців і людини є нерухомими, високодиференційованими клітинами, які у процесі розвитку втратили ядро та всі цитоплазматичні органели і пристосовані до виконання практично єдиної функції – дихальної (транспортування кисню та вуглекислого газу), що здійснюється завдяки наявності в них дихального пігменту – гемоглобіну. Кількість еритроцитів у дорослого чоловіка коливається в межах від 3,9×1012 до 6,0×1012 в 1л, а у жінок – від 3,7×1012 до 5,5×1012. Число еритроцитів у здорових людей може коливатися залежно від віку, гормонального фону, емоційного та м’язового навантаження, перебування в умовах розрідженої атмосфери тощо. Зокрема, жіночі статеві гормони гальмують розвиток еритроцитів, унаслідок чого вміст червонокрівців у крові жінок менший, ніж у чоловіків. Утворюються еритроцити в червоному кров’яному мозку із ядерних клітин – попередників, які втрачають ядро перед виходом у кров’яне русло.
Еритроцити (erythrocytus), або червонокрівці (Рис.2.1), у ссавців і людини є нерухомими, високодиференційованими клітинами, які у процесі розвитку втратили ядро та всі цитоплазматичні органели і пристосовані до виконання практично єдиної функції – дихальної (транспортування кисню та вуглекислого газу), що здійснюється завдяки наявності в них дихального пігменту – гемоглобіну. Кількість еритроцитів у дорослого чоловіка коливається в межах від 3,9×1012 до 6,0×1012 в 1л, а у жінок – від 3,7×1012 до 5,5×1012. Число еритроцитів у здорових людей може коливатися залежно від віку, гормонального фону, емоційного та м’язового навантаження, перебування в умовах розрідженої атмосфери тощо. Зокрема, жіночі статеві гормони гальмують розвиток еритроцитів, унаслідок чого вміст червонокрівців у крові жінок менший, ніж у чоловіків. Утворюються еритроцити в червоному кров’яному мозку із ядерних клітин – попередників, які втрачають ядро перед виходом у кров’яне русло.
|
 Форма і будова. Еритроцити у людини і ссавців здебільшого мають форму двоввігнутих дисків, їх називають дискоцитами. У нормі дискоцити становлять 80% від загальної кількості еритроцитів. Трапляються і інші форми еритроцитів (Рис.2.2) – планоцити (мають пласку поверхню), сфероцити (кулясті), стоматоцити (куполоподібні), ехіноцити (мають шипи) тощо. Така різноманітність форм у нормі позначається терміном фізіологічний пойкілоцитоз (з грец. «пойкілос» − різноманітний, «цитос» − клітина). Форму еритроцитів підтримують бета-сіалоглікопротеїн в еритроцитарній мембрані та спеціальний каркас, побудований з білка спектрину, що зсередини прилягає до плазмолеми і пов’язаний з нею іншим білком – анкеритом.
Форма і будова. Еритроцити у людини і ссавців здебільшого мають форму двоввігнутих дисків, їх називають дискоцитами. У нормі дискоцити становлять 80% від загальної кількості еритроцитів. Трапляються і інші форми еритроцитів (Рис.2.2) – планоцити (мають пласку поверхню), сфероцити (кулясті), стоматоцити (куполоподібні), ехіноцити (мають шипи) тощо. Така різноманітність форм у нормі позначається терміном фізіологічний пойкілоцитоз (з грец. «пойкілос» − різноманітний, «цитос» − клітина). Форму еритроцитів підтримують бета-сіалоглікопротеїн в еритроцитарній мембрані та спеціальний каркас, побудований з білка спектрину, що зсередини прилягає до плазмолеми і пов’язаний з нею іншим білком – анкеритом.
|
 центрі – до 1 мкм (Рис 2.3.). Поверхня окремого еритроцита у людини приблизно дорівнює 125 мкм2, а об’єм – 90 мкм3. Якщо рахувати, що середня кількість крові у людини 5,5 л, то загальна поверхня еритроцитів, які циркулюють у крові, складає близько 3500 – 3700 м2. Така форма клітин забезпечує збільшення їх поверхні і прискорює насичення гемоглобіну киснем. В умовах норми 75% усіх еритроцитів мають вищеназвані норми.
центрі – до 1 мкм (Рис 2.3.). Поверхня окремого еритроцита у людини приблизно дорівнює 125 мкм2, а об’єм – 90 мкм3. Якщо рахувати, що середня кількість крові у людини 5,5 л, то загальна поверхня еритроцитів, які циркулюють у крові, складає близько 3500 – 3700 м2. Така форма клітин забезпечує збільшення їх поверхні і прискорює насичення гемоглобіну киснем. В умовах норми 75% усіх еритроцитів мають вищеназвані норми.
Плазмолема еритроцитів під електронним мікроскопом має товщину близько 20 нм. На її зовнішній поверхні розташовані фосфоліпіди, сіалова кислота, антигенні олігосахариди (зумовлюють групову належність еритроцитів), адсорбовані протеїни, на внутрішній поверхні – гліколітичні ферменти, натрій- та калій-АТФази, глікопротеїни, гемоглобін. Оболонка еритроцита – це напівпроникна мембрана, яка забезпечує активне перенесення через мембрану іонів натрію та калію, О2 та СО2 та інших речовин. Усередині еритроцита розташований електронно-щільний вміст – багаточисельні гранули гемоглобіну розмірами 4 – 5 нм.
За хімічним складом еритроцити мають 60% воли та 40% сухого залишку. 95% сухого залишку складає гемоглобін і лише 5% − інші речовини. Таким чином, гемоглобін складає одну третину загальної маси еритроцита. Гемоглобін – це складний білок, побудований з білкової частини – глобіну та небілкової групи – гему, до складу якого входить залізо (Рис. 2.4).

Рис. 2.4 Структура молекули гемоглобіну
Гемоглобін є пігментом, який надає крові червоного кольору. Він здатний легко приєднувати кисень, утворюючи в легенях нестійку сполуку – оксигемоглобін, який легко розпадається і віддає кисень тканинам Звільнений від кисню гемоглобін відразу частково приєднує вуглекислий газ (продукт розпаду речовин у клітинах), утворюючи карбгемоглобін, але більша частина вуглекислоти переноситься плазмою крові. Гемоглобін також утворює сполуку з чадним газом (СО), яка має назву карбоксигемоглобіну. Спорідненість гемоглобіну із чадним газом в 300 разів вища, ніж з киснем, тому в атмосфері із значним вмістом чадного газу гемоглобін стає заблокованим, недоступним для кисню, і організм у такому випадку гине від задухи (нестачі кисню).
У людини є два типи гемоглобіну – HbA, характерний для дорослих, HbF, характерний для плоду. У дорослого HbA становить 98% і лише 2% становить HbF. У крові новонародженої дитини міститься 80% HbF та лише 20% HbА. Ряд захворювань крові (так звані гемоглобінози, або гемоглобінопатії) супроводжуються появою у крові інших типів гемоглобінів, білкова частина яких порівняно з HbА та HbF характеризується іншим набором амінокислот.
Основний шлях обміну енергії в еритроцитах – гліколіз, в процесі якого відбувається утворення АТФ та НАДН2. Енергія гліколізу забезпечує активний транспорт катіонів через плазмолему, підтримання оптимального співвідношення калію та натрію в плазмі крові та еритроцитах, а також підтримання форми та збереження цілісності мембрани еритроцита. НАДН2 підтримує активний функціональний стан гемоглобіну, запобігаючи його окисленню в метгемоглобін.
В еритроцитах також присутні ферменти пентозо-фосфатного циклу. Наявність заліза у складі гемоглобіну забезпечує жовте забарвлення еритроцитів, а у великій кількості еритроцити надають крові червоного забарвлення.
Отже, еритроцити забезпечують підтримання рН крові (гемоглобін і оксигемоглобін складають одну з буферних систем крові); підтримання іонного гомеостазу за рахунок обміну іонами між плазмою та еритроцитами; участь у водному і сольовому обміні; адсорбцію токсинів, в тому числі продуктів розпадання білка, що зменшує їх концентрацію в плазмі крові і перешкоджає переходу в тканини; участь у ферментативних процесах, у транспорті поживних речовин – глюкози, амінокислот.
Середній термін життя еритроцитів людини – 120 діб. Беручи до уваги загальне число еритроцитів в організмі та середню тривалість їхнього життя, можна підрахувати, що упродовж доби руйнується 200 мільйонів еритроцитів і стільки ж утворюється їм на зміну. У крові, таким чином, можна знайти різні за віком еритроцити: молоді, функціонально зрілі і такі, що старіють. Молоді форми еритроцитів носять назву ретикулоцитів. Вони не повністю насичені гемоглобіном, їм властива поліхроматофілія (здатність забарвлюватися як кислими, так і основними барвниками). У своїй цитоплазмі ретикулоцити містять сітчасту структуру (звідси походить назва цих клітин), яку можна виявити прижиттєвим фарбуванням мазка крові діамант-крезиловим синім. Електронно-мікроскопічно доведено, що сітчаста структура в цитоплазмі ретикулоцита – це залишки гранулярної ендоплазматичної сітки та вільних рибосом, на яких продовжується синтез гемоглобіну, а також мітохондрій. У нормі кількість ретикулоцитів становить 1 – 5% від загальної кількості еритроцитів. Збільшення їх кількості є діагностичною ознакою посиленого кровотворення.
При старінні еритроцитів спостерігається зменшення активності різноманітних ферментів, зокрема, гексокінази, глюкозо-6-фосфатдегідрогенази, що призводить до зниження інтенсивності гліколізу і реакцій пентозного циклу. Має місце також зменшення вмісту ліпідів та АТФ і, внаслідок цього, збільшення чутливості до осмотичного лізису та механічного впливу. В результаті порушення проникності мембрани спостерігається вивільнення іонів калію в плазму і збільшення в еритроцитах вмісту натрію. В еритроцитах, що старіють відмічається порушення газообмінної функції внаслідок зменшення здатності до відновлення метгемоглобіну в гемоглобін.
Лейкоцити, або білі кров’яні тільця. На відміну від еритроцитів, лейкоцити мають ядро і всі цитоплазматичні органели, не мають пігменту, здатні до виходу із судин та активного пересування шляхом утворення псевдоподій; виконують захисну функцію; основний термін життя проводять поза кровоносними судинами. У дорослої людини в 1 л крові міститься від 4,0×109 до 10,0×109 лейкоцитів.
Основною функцією лейкоцитів є здійснення імунних реакцій організму: вони руйнують різноманітні генетично чужорідні агенти, які потрапляють до організму, а також руйнують власні відмерлі або змінені (онко-) клітини. Виконують захисну функцію шляхом фагоцитозу, цитотоксичної дії або вироблення антитіл.
Лейкоцити – група клітин, які відрізняються одна від одної за своїм походженням, морфологічними (величина клітин, будова ядра, наявність гранул з фізіологічно активними речовинами в цитоплазмі), цитохімічними і функціональними властивостями. У нормі у здорової дорослої людини в 1 мм3 крові міститься 5000 – 7000 лейкоцитів.
Лейкоцити, в залежності від наявності специфічних гранул в їх цитоплазмі, поділяються на гранулоцити (нейтрофіли, базофіли, еозинофіли) та агранулоцити (лімфоцити, моноцити).
|
 Нейтрофіли– лейкоцити округлі клітини, діаметром 10 – 15 мкм (Рис. 2.5). Вони відрізняються формами ядер, тому їх часто називають поліморфно-ядерними лейкоцитами. За формою ядра нейтрофіли розділяються на юні, палочкоядерні і сегментовані. Юні і паличкоядерні клітини є незрілими формами нейтрофілів. Цитоплазма нейтрофілів блідо-рожева з обширною, нерівномірною і дрібною зернистістю, забарвленою в рожево-синій або фіолетовий колір. Ядро темно-фіолетове паличкоподібне, або сегментоване (2 – 5 сегментів). Нейтрофіли – найчисленніша група лейкоцитів периферійної крові. Вони складають 48 – 78 % від загального числа лейкоцитів.
Нейтрофіли– лейкоцити округлі клітини, діаметром 10 – 15 мкм (Рис. 2.5). Вони відрізняються формами ядер, тому їх часто називають поліморфно-ядерними лейкоцитами. За формою ядра нейтрофіли розділяються на юні, палочкоядерні і сегментовані. Юні і паличкоядерні клітини є незрілими формами нейтрофілів. Цитоплазма нейтрофілів блідо-рожева з обширною, нерівномірною і дрібною зернистістю, забарвленою в рожево-синій або фіолетовий колір. Ядро темно-фіолетове паличкоподібне, або сегментоване (2 – 5 сегментів). Нейтрофіли – найчисленніша група лейкоцитів периферійної крові. Вони складають 48 – 78 % від загального числа лейкоцитів.
Основна ефекторна функція нейтрофілів – це фагоцитоз, який вони можуть здійснювати тільки один раз, після чого гинуть.
Нейтрофіли не мають специфічності і здатні знищувати бактерії, клітини крові, продукти розпаду тканин, неорганічні частки.
Плазма нейтрофілів містить гранули. Первинні гранули є лізосомами і містять лізоцим, кислі гідролази і мієлопероксидазу. Вторинні гранули називають специфічними. Поряд з лізоцимами вони містять лактоферин і антибіотичні білки, що руйнують клітини мікроорганізмів (дефензини, сепроцидини, кателицидини, білок, який сприяє підвищенню проникності клітинної стінки бактерій).
|
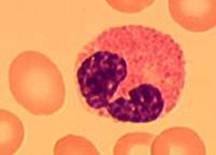 Еозинофіли – округлі клітини, діаметром 12 – 17 мкм. В цитоплазмі містять близько 200 гранул.. Ядро рихле, широке, складається з 2-3 сегментів, забарвлене у фіолетовий колір (Рис 2.6). Еозинофіли складають 1 – 5% усіх лейкоцитів. Це – спеціалізована популяція лейкоцитів, здатних вражати великих позаклітинних паразитів (гельмінтів), які не піддаються фагоцитозу. Токсикогенність еозинофілів обумовлена реакцією дегрануляції: багаточисленних гранул, що містять токсичні речовини. Зливаючись з цитоплазматичною мембраною, вміст гранул вивільняється в позаклітинне середовище еозинофілу.
Еозинофіли – округлі клітини, діаметром 12 – 17 мкм. В цитоплазмі містять близько 200 гранул.. Ядро рихле, широке, складається з 2-3 сегментів, забарвлене у фіолетовий колір (Рис 2.6). Еозинофіли складають 1 – 5% усіх лейкоцитів. Це – спеціалізована популяція лейкоцитів, здатних вражати великих позаклітинних паразитів (гельмінтів), які не піддаються фагоцитозу. Токсикогенність еозинофілів обумовлена реакцією дегрануляції: багаточисленних гранул, що містять токсичні речовини. Зливаючись з цитоплазматичною мембраною, вміст гранул вивільняється в позаклітинне середовище еозинофілу.
Крім того, еозинофіли утворюють токсичні метаболіти кисню. Обидва механізми складають основу противогельмінтного імунітету.
Оксифілія гранул еозинофілів зумовлена наявністю в них головного лужного білка (МВР – від англ. major basic protein), багатого на аргінін. Серед складників специфічних гранул ацидофілів переважають гідролази та пероксидази, тому ці гранули вважають різновидом лізосом або пероксисом. Крім того, гранули еозинофілів містять фермент гістаміназу, а лізоцим і фагоцитин у них відсутні. Органели цитоплазми розвинені слабко.
Еозинофіли у червоному кістковому мозку проходять ті ж стадії розвитку, що й нейтрофіли, тобто існують юні, паличкоядерні та сегментоядерні еозинофіли. Але, оскільки вміст цих клітин у крові невеликий, юні і паличкоядерні форми еозинофілів трапляються дуже рідко і під час підрахунків не враховуються. Ядро в сегментоядерних еозинофілах найчастіше складається з двох, рідше – з трьох сегментів. Сегменти більші, ніж у нейтрофілів. Структура ядра ніжніша, сегменти більш правильної форми. До складу ядер еозинофілів в основному входить гетерохроматин, ядерця не помітні.
Еозинофільні лейкоцити рухомі, здатні до фагоцитозу, однак їхня фагоцитарна активність нижча, ніж у нейтрофілів. Вони приймають участь у захисних реакціях організму на сторонній білок, в алергійних та анафілактичних реакціях. Еозинофіли не здатні до синтезу гістаміну, однак здатні його накопичувати, фагоцитуючи гістамінвмісні гранули, які виділяють базофіли та тічні клітини. А також здатні адсорбувати гістамін на цитолемі, що містить рецептори до гістаміну.
Базофіли – округлі клітини, діаметром 9– 12 мкм. Становлять 0 – 1% від загальної кількості лейкоцитів. Цитоплазма заповнена крупними, кулястими або полігональними базофільними гранулами. Специфічна зернистість фарбується за Романовським інтенсивно в пурпурово-фіолетовий колір.. Розміри гранул 0,5 – 1,2 мкм (Рис 2.7). Метахромазія гранул зумовлена наявністю в них кислого глюкозаміноглікану гепарину. Крім того, в гранулах міститься гістамін, серотонін, пероксидаза, кисла фосфатаза, а також фермент синтезу гістаміну – гістидиндекарбоксилаза. Ядро базофілів не має певної форми (сегментоване, богоподібне, рідше – сферичне тощо), розташоване в центрі клітини, порівняно бідне на гетерохроматин. Ядро фарбується менш інтенсивно, ніж зернистість, унаслідок чого остання прикриває та маскує його. В цитоплазмі виявляються всі види основних органел.
Базофіли – малорухомі клітини, майже не здатні до фагоцитозу. Їхня функція полягає у метаболізмі гістаміну та гепарину.
Гепарин є нативним антикоагулянтом, тому базофіли, так як і тучні клітини сполучної тканини, беруть участь у регуляції процесу згортання крові та проникності судин. Гістамін зумовлює різке розширення судин, появу набряків тощо. Такі явища виникають у разі дегрануляції базофілів, які, таким чином, приймають участь в алергійних реакціях.
Гепарин – це сульфатований глікозаміноглікан, який уперше було виділено з печінки (цим зумовлена його назва) і який запобігає згортанню крові. Виявлено, що тканинні базофіли синтезують гепарин у комплексі Гольджі. Вони можуть втрачати свої гранули (процес дегрануляції), і тоді гепарин виділяється у міжклітинну речовину. Гепарин знижує її проникність, має протизапальну дію, є антикоагулянтом. Крім того, гепарин стимулює активність ферменту ліпопротеїнліпази.
Гістамін синтезується у тканинних базофілах за участі гістидиндекарбоксилази (маркерний фермент цих клітин), яка здійснює перетворення гістидину у гістамін, що діє на гладкі м’язи, спричиняючи їхнє скорочення, а також сприяє виходу плазми з венул і капілярів за рахунок розширення і підвищення проникності їх стінок. Унаслідок виходу плазми у пухкій сполучній тканині під епідермісом утворюються пухирці. Цей симптом отримав назву кропивниці.
Описану дію гістаміну можна спостерігати під час анафілактичного шоку або алергії. Розвиток цих процесів та участь у них тканинних базофілів пояснюється так. У відповідь на проникнення до організму деяких антигенів, що носять назву алергенів, утворюються специфічні антитіла, які належать до класу імуноглобулінів Е (IgE).
Тканинні базофіли, як і базофільні лейкоцити, мають рецептори до антитіл цього типу і зв’язують їх так, що варіабельні ділянки молекул імуноглобулінів залишаються вільними. У разі повторного введення останній сполучається з антитілами на поверхні тканинних базофілів. Після утворення комплексу антиген-антитіло гістамін вивільняється з гранул цих клітин. Симптоми алергії або анафілаксії можна усунути введенням антигістамінних препаратів. За нормальних умов такі реакції гіперчутливості, які відбуваються за участю тканинних базофілів, мають тенденцію до самообмеження внаслідок виділення цими клітинами хемотаксичного фактору залучення еозинофілів. Ферменти еозинофілів гістаміназа, арилсульфатаза руйнують речовини, які вивільняють тканинні базофіли під час імунних реакцій.
Тканинні базофіли мають багато назв, які доцільно навести, щоб допомогти орієнтуватися в літературі: мастоцити, лаброцити, тучні клітини. Останню назву дав цим клітинам П. Еріх, який у 1877 р. вперше описав клітини, що були переповнені гранулами, ніби «об’їлися» ними. Ця назва дуже поширена в літературі. Назва «тканинні базофіли» свідчить про те, що клітини мають зернистість подібну до гранул базофільних лейкоцитів крові. Тканинні базофіли часто локалізують уздовж кровоносних судин мікроциркуляторного русла. Велика кількість цих клітин зустрічається у стінці органів травного каналу, в матці, молочній залозі, тимусі, мигдаликах.
Форма тканинних базофілів різноманітна, так само як і розміри. Вони бувають круглі, овальні, з широкими відростками. Розміри коливаються від 10 – 20 до 35 і навіть до 100 мкм. Ядра порівняно невеликі, круглі, звичайної будови. У цитоплазмі міститься велика кількість мітохондрій, небагато елементів гранулярної, а також агранулярної ендоплазматичної сітки; добре розвинений комплекс Гольджі.
Головна особливість цих клітин – наявність великої кількості характерних гранул розмірами 0,2 – 0,8 мкм, кожна з яких оточена мембраною. За електронно-мікроскопічною будовою гранули тканинних базофілів людини кристалоїдні або пластинчасті (спостерігаються видові відмінності структури гранул). Гранули містять кілька речовин, що мають фізіологічне значення. Першою з таких речовин є гепарин, який становить 30% вмісту гранул. Друга речовина – гістамін, який становить 10% їх вмісту. Матрикс гранули складається з білка та гепарину, які формують стабільну сітку; до неї іонними зв’язками приєднаний гістамін. До складу гранул також входять хондроїтидинсульфат, гіалуронова кислота, у деяких тварин (але не у людини) знайдено і серотонін.
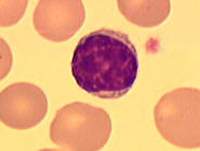
Лімфоцити– клітини імунної системи (імунокомпетентні клітини), вид лейкоцитів, які відповідають за набутий імунітет.
Лімфоцити у крові дорослих становлять 20 – 40% від загальної кількості лейкоцитів. Залежно від розмірів на рівні світлової мікроскопії розрізняють три види лімфоцитів – малі (діаметром 4,5 – 7 мкм, становлять за кількістю 2/3 від усіх лімфоцитів крові), середні (діаметром 7 – 10 мкм і становлять 1/3 всіх лімфоцитів) та великі (діаметром понад 10 мкм), у крові дорослих не зустрічаються, їх можна знайти лише у лімфі грудної протоки.
|
|
За даними електронної мікроскопії серед лімфоцитів розрізняють чотири типи клітин:
1. Малі світлі лімфоцити. Їх найбільше – 70 – 75%. Вони мають світлу цитоплазму з невеликою кількістю вільних рибосом, містять також усі інші органели.
|
3. Середні лімфоцити. Їх 10 – 12%. Хроматин пухкий, добре видно ядерце. У цитоплазмі містяться практично всі органели. В цитоплазмі розташовані подовжені канальці ендоплазматичної сітки, є елементи агранулярної сітки, вільні рибосоми і полісоми. Центрисома та комплекс Гольджі зазвичай розташовані поряд з областю інвагінації ядерної мембрани, мітохондрії дрібні, лізосоми зустрічаються у невеликій кількості.
4. Плазмоцити або лімфоплазмоцити. Вони становлять 1 – 2%. Характерна їхня ознака – концентрично розташовані навколо ядра канальці ендоплазматичної сітки. Плазмоцити мають розміри 7 – 10 мкм, хоча можуть бути дещо більшими. Форма їх округла або багатокутна, якщо вони прилягають одна до одної. Ядро невелике, кругле, розташоване ексцентрично, містить переважно конденсований хроматин, грудочки якого утворюють характерний для плазмоцита малюнок – колеса зі спицями або цифри на циферблаті годинника. Ультраструктура цих клітин характеризується наявністю у цитоплазмі добре розвиненої гранулярної ендоплазматичної сітки, що розташована концентрично і займає більшу частину клітини. Велика кількість рибосом (РНК) зумовлює базофілію цитоплазми. В цитоплазмі локалізовані центріолі, оточені цистернами комплексу Гольджі. У цистернах гранулярної ендоплазматичної сітки плазмоцитів відбувається синтез імуноглобулінів (антитіл). Частина вуглеводного компонента імуноглобулінів синтезується у комплексі Гольджі. Ця органела, яка досить добре розвинена у плазмоцитах, відповідає також за секрецію синтезованих імуноглобулінів за межі клітини; далі вони потрапляють через лімфу у кров.
Таким чином плазмоцити забезпечують гуморальний імунітет, тобто вироблення специфічних білків – імуноглобулінів (антитіл), реагуючи на проникнення в організм антигену, який буде знешкоджуватися антитілами. Походять плазматичні клітини зі стовбурової кровотворної клітини (через стадію В-лімфоцитів). Плазматичні клітини здебільшого зустрічаються у пухкій сполучній тканині власної пластинки слизової оболонки кишки та дихальних шляхів, у лімфатичних вузлах, селезінці, у сполучній тканині різних залоз.
Моноцити – округлі клітини, розмір діаметр 10 – 20 мкм, ядро світло-фіолетового або бузкового кольору, у вигляді боба, метелика, вісімки або гриба, займає велику частину клітини (Рис. 2.9). Моноцити в нормі складають 2 – 8% від загальної кількості лейкоцитів.
|
 З крові моноцити мігрують в тканини, де перетворюються в тканинні макрофаги. За діаметром ці клітини найбільші серед білокрівців, особливо на мазках, унаслідок сильного розпластування їх на склі (діаметр при цьому становить 18 – 20 мкм). У краплі свіжої крові їх розміри значно менші (10 – 12 мкм). Цитоплазма фарбується базофільно, але не так яскраво, як у лімфоцита, а має димчасто-сірий відтінок. У цитоплазмі знаходяться усі органели, численні лізосоми. На відміну від гранулоцитів моноцити не містять цитоплазматичних гранул, але іноді містить у великій кількості дрібну азурофільну зернистість.
З крові моноцити мігрують в тканини, де перетворюються в тканинні макрофаги. За діаметром ці клітини найбільші серед білокрівців, особливо на мазках, унаслідок сильного розпластування їх на склі (діаметр при цьому становить 18 – 20 мкм). У краплі свіжої крові їх розміри значно менші (10 – 12 мкм). Цитоплазма фарбується базофільно, але не так яскраво, як у лімфоцита, а має димчасто-сірий відтінок. У цитоплазмі знаходяться усі органели, численні лізосоми. На відміну від гранулоцитів моноцити не містять цитоплазматичних гранул, але іноді містить у великій кількості дрібну азурофільну зернистість.
Моноцити рухомі, здатні до фагоцитозу і піноцитозу. Їхня здатність до адгезії обумовлена фагоцитарною активністю. Розрізняють незрілі моноцити, що циркулюють у крові, та зрілі їх похідні, до яких належать тканинні макрофаги та дендритні клітини.
Незрілі моноцити мають округлу форму і невеликі розміри, а тканинні моноцити, як правило, – амебоїдну форму з псевдоподіями та дендритами, утвореними за рахунок вигинів плазматичної мембрани.
Моноцити перебувають у кров’яному руслі недовго – від 36 до 104 год, після чого виходять із судин і в тканинах перетворюються на макрофаги-гістоцити, які є кінцевою стадією диференціації цих клітин.
Тромбоцити, або кров’яні пластинки мають вид дрібних безбарвних тілець округлої, овальної, веретеноподібної або неправильної форми. Тромбоцити – це без’ядерні фрагменти цитоплазми гігантських колоній кісткового мозку – мегакаріоцитів, розміром 2 – 3 мкм. Кількість тромбоцитів 200 – 400×109 в 1 л крові. Підрахувати ці формені елементи важко через здатність їх склеюватися в конгломерати.
Мегакаріоцит – найбільша клітина червоного кісткового мозку, її розміри від 50 – 70 до 100 мкм. Ядро поліморфне, фрагментоване, із заглибленнями і вирізами, дуже химерної (примхливої) форми, структура його грубо сітчаста, ядерець немає. Цитоплазма базофільна, фарбується у фіолетовий колір, містить азурофільну зернистість. Клітина нечітко відмежована від навколишнього середовища. Цитоплазма сегментується на невеликі фрагменти, які відокремлюються від клітини, перетворюючись на кров’яні пластинки (тромбоцити). З одного мегакаріоцита утворюється 3 – 4×103 тромбоцитів. Особливість мегакаріоцитів полягає у тому, що ці клітини є поліплоїдними, число хромосомних наборів у них може досягати 32 – 64. Поліплоїдізація цих клітин зумовлена тим, що на шляху їхнього утворення з мегакаріобластів поділи не відбуваються, а здійснюються 4 – 5 ендомітозів, у результаті чого збільшується об’єм ядра, і цитоплазми.
Кожна кров’яна пластинка (тромбоцит) складається з гіаломера, що є її основою, та грануломера (або хромомера), який має вигляд зерняток у центрі пластинки. Грануломер не містить у собі ДНК. Ззовні кров’яні пластинки оточені плазмолемою. У гіаломері міститься крайовий пучок мікротрубочок, який допомагає тромбоциту утримувати форму. Тут також містяться актинові і міозинові мікрофіламенти. Крім того, в грануломері наявні мітохондрії та різні кількості гранул глікогену. У складі грануломера електронна мікроскопія виявила два типи гранул: щільні, темні альфа-гранули, хімічний склад яких недостатньо вивчений, і серотонінові гранули. У гарнуломері є також зерна глікогену і мітохондрії. Кров’яні пластинки мають відростки різних розмірів і товщини (так звані вусики). Цими відростками пластинки зчіплюються одна з одною під час згортання крові. Відсутність вусиків у тромбоцитах супроводжується порушенням процесів згортання крові.
|
|
 також виділяють речовини, що спричиняють звуження судини у разі її ушкодження та зменшення проникності судинної стінки.
також виділяють речовини, що спричиняють звуження судини у разі її ушкодження та зменшення проникності судинної стінки.
|
Гемограма. Лейкоцитарна формула.У крові здорової людини формені елементи знаходяться у певних кількісних співвідношеннях, що називають гемограмою. Відсоткові співвідношення різних видів лейкоцитів у мазку периферійної крові складають лейкоцитарну формулу (Табл.2.1).
Таблиця 2.1
Читайте також:
- I. Загальні збори АТ
- I. ЗАГАЛЬНІ МЕТОДИЧНІ ВКАЗІВКИ
- I. Загальні положення
- II. ЗАГАЛЬНІ ПОЛОЖЕННЯ.
- IX. Відомості про військовий облік
- IX. Відомості про військовий облік
- V Практично всі психічні процеси роблять свій внесок в специфіку організації свідомості та самосвідомості.
- XXXIII. ЗАГАЛЬНІ ПРОФЕСІЇ (У ВСІХ ГАЛУЗЯХ ГОСПОДАРСТВА)
- А) загальні критерії
- Білковий обмін: загальні відомості
- Біографічні відомості
- Боротьба з проявами національної самосвідомості
| <== попередня сторінка | | | наступна сторінка ==> |
| Підрахунок лейкоцитарної формули. | | | Показники гемограми та лейкоцитарної формули здорової людини |
|
Не знайшли потрібну інформацію? Скористайтесь пошуком google: |
© studopedia.com.ua При використанні або копіюванні матеріалів пряме посилання на сайт обов'язкове. |