РЕЗОЛЮЦІЯ: Громадського обговорення навчальної програми статевого виховання
ЧОМУ ФОНД ОЛЕНИ ПІНЧУК І МОЗ УКРАЇНИ ПРОПАГУЮТЬ "СЕКСУАЛЬНІ УРОКИ"
ЕКЗИСТЕНЦІЙНО-ПСИХОЛОГІЧНІ ОСНОВИ ПОРУШЕННЯ СТАТЕВОЇ ІДЕНТИЧНОСТІ ПІДЛІТКІВ
Батьківський, громадянський рух в Україні закликає МОН зупинити тотальну сексуалізацію дітей і підлітків
Відкрите звернення Міністру освіти й науки України - Гриневич Лілії Михайлівні
Представництво українського жіноцтва в ООН: низький рівень культури спілкування в соціальних мережах
Гендерна антидискримінаційна експертиза може зробити нас моральними рабами
ЛІВИЙ МАРКСИЗМ У НОВИХ ПІДРУЧНИКАХ ДЛЯ ШКОЛЯРІВ
ВІДКРИТА ЗАЯВА на підтримку позиції Ганни Турчинової та права кожної людини на свободу думки, світогляду та вираження поглядів
- Гідрологія і Гідрометрія
- Господарське право
- Економіка будівництва
- Економіка природокористування
- Економічна теорія
- Земельне право
- Історія України
- Кримінально виконавче право
- Медична радіологія
- Методи аналізу
- Міжнародне приватне право
- Міжнародний маркетинг
- Основи екології
- Предмет Політологія
- Соціальне страхування
- Технічні засоби організації дорожнього руху
- Товарознавство продовольчих товарів
Тлумачний словник
Авто
Автоматизація
Архітектура
Астрономія
Аудит
Біологія
Будівництво
Бухгалтерія
Винахідництво
Виробництво
Військова справа
Генетика
Географія
Геологія
Господарство
Держава
Дім
Екологія
Економетрика
Економіка
Електроніка
Журналістика та ЗМІ
Зв'язок
Іноземні мови
Інформатика
Історія
Комп'ютери
Креслення
Кулінарія
Культура
Лексикологія
Література
Логіка
Маркетинг
Математика
Машинобудування
Медицина
Менеджмент
Метали і Зварювання
Механіка
Мистецтво
Музика
Населення
Освіта
Охорона безпеки життя
Охорона Праці
Педагогіка
Політика
Право
Програмування
Промисловість
Психологія
Радіо
Регилия
Соціологія
Спорт
Стандартизація
Технології
Торгівля
Туризм
Фізика
Фізіологія
Філософія
Фінанси
Хімія
Юриспунденкция
Спряження збудження та скорочення. Механізми м’язового скорочення.
 Термін “спряження збудження із скороченням” означає взаєзв’язок збудження в скелетних м’язах (виникнення та поширення ПД по мембрані волокна) та його скорочення, тобто актоміозинової взаємодії.
Термін “спряження збудження із скороченням” означає взаєзв’язок збудження в скелетних м’язах (виникнення та поширення ПД по мембрані волокна) та його скорочення, тобто актоміозинової взаємодії.
В стані спокою взаємодії актинових та міозинових протофібрил немає тому, що активні центри актину заблоковані регуляторними білками тропоніном та тропоміозином. Знімають блокаду іони Са2+, концентрація яких в саркоплазмі в стані спокою низька 10–8 ммоль/л.
ПД, що виникають в постсинаптичній мембрані поширюються вздовж всієї довжини мембрани м’язового волокна, у тому числі і по мембрані Т-трубочок (це вгинання зовнішньої мембрани м’язових клітин всередину волокна).
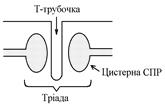 Безпосередньо поруч з Т-трубочками розташовані цистерни саркоплазматичного ретикулуму (СПР – частина аграпулярної епдоплазматичної сітки). Т-трубочки з двома поруч розташованими цистернами утворюють тріади. СПР крім цистернмаютьпоздовжні трубочки, в яких локалізуються кальцієві насоси – вони активно транспортують іони – Са2+із саркоплазми в СПР і в його цистернах накопичується велика кількість іонів Са2+. При русі ПД по мембрані Т-трубочок в мембрані цистерн СПР відкриваються кальцієві канали Ù іони Са2+ по градієнту концентрації виходять з цистерн СПР у саркоплазму Ù підвищення концентрації іонів Са2+ в саркоплазмі міоцита з 10–8 до 10–5 ммоль/л Ù дифузія іонів Са2+ до протофібрил Ù взаємодія з регуляторним білком тропоніном Ù зміна третинної конформації тропоніну та тропоміозину Ù відкриття активних центрів актину Ù взаємодія головок міозину з активними центрами актину (м’язове скорочення).
Безпосередньо поруч з Т-трубочками розташовані цистерни саркоплазматичного ретикулуму (СПР – частина аграпулярної епдоплазматичної сітки). Т-трубочки з двома поруч розташованими цистернами утворюють тріади. СПР крім цистернмаютьпоздовжні трубочки, в яких локалізуються кальцієві насоси – вони активно транспортують іони – Са2+із саркоплазми в СПР і в його цистернах накопичується велика кількість іонів Са2+. При русі ПД по мембрані Т-трубочок в мембрані цистерн СПР відкриваються кальцієві канали Ù іони Са2+ по градієнту концентрації виходять з цистерн СПР у саркоплазму Ù підвищення концентрації іонів Са2+ в саркоплазмі міоцита з 10–8 до 10–5 ммоль/л Ù дифузія іонів Са2+ до протофібрил Ù взаємодія з регуляторним білком тропоніном Ù зміна третинної конформації тропоніну та тропоміозину Ù відкриття активних центрів актину Ù взаємодія головок міозину з активними центрами актину (м’язове скорочення).
В основі м’язового скорочення лежить ковзання актинових протофібрил відносно міозинових “теорія ковзання”.
 Саркомер – сруктурно-функціональний елемент скорочувального апарату скелетних м’язів. Вони утворені пучками міофібрил, які відокремлені один від одного перпендикулярними смугами – Z-лініями. До Z-ліній прикріпляються одним своїм кінцем тонкі актинові нитки. Інші кінці актинових ниток спрямовані до центру саркомера і входять у проміжки між товстими міозиновими нитками. Частина саркомера, яка примикає до Z лінії та утворена тільки актиновими протофібрилами, має назву І-дисків (ізотропних); слідом за ними розташовані А-диски (анізотропні) – частина саркомера, де має місце взаємне перекриття актинових та міозинових протофібрил. При вкороченні м’яза, в, ході його скорочення, коротшає довжина протофібрил Ù вкорочення довжини кожного саркомера. Але при цьому довжина анізотропних дисків не зменшується, а зменшується довжина ізотропних дисків. Це є наслідком ковзання актинових протофібрил відносно міозинових в напрямку до центру саркомера.
Саркомер – сруктурно-функціональний елемент скорочувального апарату скелетних м’язів. Вони утворені пучками міофібрил, які відокремлені один від одного перпендикулярними смугами – Z-лініями. До Z-ліній прикріпляються одним своїм кінцем тонкі актинові нитки. Інші кінці актинових ниток спрямовані до центру саркомера і входять у проміжки між товстими міозиновими нитками. Частина саркомера, яка примикає до Z лінії та утворена тільки актиновими протофібрилами, має назву І-дисків (ізотропних); слідом за ними розташовані А-диски (анізотропні) – частина саркомера, де має місце взаємне перекриття актинових та міозинових протофібрил. При вкороченні м’яза, в, ході його скорочення, коротшає довжина протофібрил Ù вкорочення довжини кожного саркомера. Але при цьому довжина анізотропних дисків не зменшується, а зменшується довжина ізотропних дисків. Це є наслідком ковзання актинових протофібрил відносно міозинових в напрямку до центру саркомера.
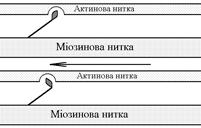 Причиною руху актинових ниток відносно міозинових (їх ковзання) є “нахил” голівки після її приєднання до активного центру актину. Велика кількість голівок, що входять до складу актоміозинових містків, одночасно роблять “нахил” Ù нитка актину переміщується до центру саркомера вздовж нитки міозину. За рахунок одного такого “нахилу” саркомер (м’яз) може вкоротитися на 1% своєї вихідної довжини. Ступінь вкорочення м’яза може досягати 50% Ù такі “нахили” голівок під час одного м’язового скорочення мають повторитися 50 разів. Це можливо, якщо актоміозинові містки після “нахилу” розпадаються (дисоціюють) Ù голівка міозину взаємодіє з наступним активним центром актину Ù наступний “нахил” Ù розпад містку і т.д. На “нахил” однієї головки міозину витрачається енергія однієї молекули АТФ. Для того, щоб відбувся розпад актоміозинового містка необхідно, щоб до голівки міозину приєдналася молекула АТФ (але енергія АТФ при цьому не витрачається).
Причиною руху актинових ниток відносно міозинових (їх ковзання) є “нахил” голівки після її приєднання до активного центру актину. Велика кількість голівок, що входять до складу актоміозинових містків, одночасно роблять “нахил” Ù нитка актину переміщується до центру саркомера вздовж нитки міозину. За рахунок одного такого “нахилу” саркомер (м’яз) може вкоротитися на 1% своєї вихідної довжини. Ступінь вкорочення м’яза може досягати 50% Ù такі “нахили” голівок під час одного м’язового скорочення мають повторитися 50 разів. Це можливо, якщо актоміозинові містки після “нахилу” розпадаються (дисоціюють) Ù голівка міозину взаємодіє з наступним активним центром актину Ù наступний “нахил” Ù розпад містку і т.д. На “нахил” однієї головки міозину витрачається енергія однієї молекули АТФ. Для того, щоб відбувся розпад актоміозинового містка необхідно, щоб до голівки міозину приєдналася молекула АТФ (але енергія АТФ при цьому не витрачається).
Для розслаблення скелетного м’язу необхідно, щоб зменшилась концентрація іонів Са2+ в саркоплазмі. Це досягається завдяки дії кальцієвих насосів повздовжніх трубочок СПР Ù активний транспорт іонів Са2+ в СПР Ù зменшення його концентрації в саркоплазмі до рівня 10–8 ммоль/л Ù від’єднання іонів Са2+ від регуляторного білка тропоніну Ù відновлення вихідної третинної структури тропоніну та тропоміозину Ù блокада активних центрів актину Ù розслаблення м’язу.
Таким чином, під час м’язового скорочення енергія АТФ витрачається:
1. На роботу натрій-калієвого насосу зовнішньої мембрани м’язового волокна (цей насос підтримує градієнт концентрації іонів К+ та Na+ поза та всередині клітини, що необхідно для підтримки на необхідному рівні ПС та для розвитку ПД на мембрані).
2. На “нахил” головки міозину.
3. На роботу кальцієвого насосу СПР Ù розслаблення м’язу.
Читайте також:
- II. МЕХАНІЗМИ ФІЗІОЛОГІЧНОЇ ДІЇ НА ОРГАНІЗМ ЛЮДИНИ.
- V Процес інтеріоризації забезпечують механізми ідентифікації, відчуження та порівняння.
- Альдостерон та механізми ренін-ангіотензину
- Анатомо-фізіологічні механізми безпеки і захисту людини від впливів негативно діючих факторів.
- Асинхронний генератор з конденсаторним збудженням.
- Біоелектричні явища і збудження в тканинах.
- БІОХІМІЯ М’ЯЗІВ І М’ЯЗОВОГО СКОРОЧЕННЯ
- Будова та функції нервово-м’язового синапсу.
- Вибір елементів пуско-гальмівного реостата для двигунів послідовного збудження
- Вибір елементів пускового (гальмівного) реостата для двигунів змішаного збудження
- Види атак на механізми та протоколи автентифікації
- Види центрального гальмування. Механізми розвитку пре- та постсинаптичного гальмування.
| <== попередня сторінка | | | наступна сторінка ==> |
| Механізми проведення збудження через нервово-м’язовий синапс. Медіатор, мембранні циторецептори та блокатори нервово-м’язових синапсів. | | | Види м’язових скорочень: одинокі та тетанічні; ізотонічні та ізометричні. |
|
Не знайшли потрібну інформацію? Скористайтесь пошуком google: |
© studopedia.com.ua При використанні або копіюванні матеріалів пряме посилання на сайт обов'язкове. |