РЕЗОЛЮЦІЯ: Громадського обговорення навчальної програми статевого виховання
ЧОМУ ФОНД ОЛЕНИ ПІНЧУК І МОЗ УКРАЇНИ ПРОПАГУЮТЬ "СЕКСУАЛЬНІ УРОКИ"
ЕКЗИСТЕНЦІЙНО-ПСИХОЛОГІЧНІ ОСНОВИ ПОРУШЕННЯ СТАТЕВОЇ ІДЕНТИЧНОСТІ ПІДЛІТКІВ
Батьківський, громадянський рух в Україні закликає МОН зупинити тотальну сексуалізацію дітей і підлітків
Відкрите звернення Міністру освіти й науки України - Гриневич Лілії Михайлівні
Представництво українського жіноцтва в ООН: низький рівень культури спілкування в соціальних мережах
Гендерна антидискримінаційна експертиза може зробити нас моральними рабами
ЛІВИЙ МАРКСИЗМ У НОВИХ ПІДРУЧНИКАХ ДЛЯ ШКОЛЯРІВ
ВІДКРИТА ЗАЯВА на підтримку позиції Ганни Турчинової та права кожної людини на свободу думки, світогляду та вираження поглядів
- Гідрологія і Гідрометрія
- Господарське право
- Економіка будівництва
- Економіка природокористування
- Економічна теорія
- Земельне право
- Історія України
- Кримінально виконавче право
- Медична радіологія
- Методи аналізу
- Міжнародне приватне право
- Міжнародний маркетинг
- Основи екології
- Предмет Політологія
- Соціальне страхування
- Технічні засоби організації дорожнього руху
- Товарознавство продовольчих товарів
Тлумачний словник
Авто
Автоматизація
Архітектура
Астрономія
Аудит
Біологія
Будівництво
Бухгалтерія
Винахідництво
Виробництво
Військова справа
Генетика
Географія
Геологія
Господарство
Держава
Дім
Екологія
Економетрика
Економіка
Електроніка
Журналістика та ЗМІ
Зв'язок
Іноземні мови
Інформатика
Історія
Комп'ютери
Креслення
Кулінарія
Культура
Лексикологія
Література
Логіка
Маркетинг
Математика
Машинобудування
Медицина
Менеджмент
Метали і Зварювання
Механіка
Мистецтво
Музика
Населення
Освіта
Охорона безпеки життя
Охорона Праці
Педагогіка
Політика
Право
Програмування
Промисловість
Психологія
Радіо
Регилия
Соціологія
Спорт
Стандартизація
Технології
Торгівля
Туризм
Фізика
Фізіологія
Філософія
Фінанси
Хімія
Юриспунденкция
Структура теломер
Нуклеотидна структура теломерних послідовностей ДНК нам вже відома. Вищі рівні організації утворюються за рахунок специфічних білків.
Мабуть, ці білки, на відміну від звичайних гістонів, не утворюють нуклеосомні глобули. Нуклеосомна структура в достатньо коротких теломерах не виявлена. Хоча довгі теломери мишей мають нуклеосомну організацію.
Найвідоміші серед теломерних білків - білок Rар1 (у дріжджів) і його аналог білок ТRF1 (у ссавців).
Очевидно, завдяки цим білкам, теломери мають щільну упаковку, тобто відносяться до фракції гетерохроматина.
У свою чергу, така структура робить теломери дуже стабільними. Зокрема, теломерні повтори недоступні для теломерази упродовж більшої частини клітинного цикла. Очевидно, в S-фазі у відповідь на деякий сигнал білок ТRF1 дисоціює від теломери - починається її подовження. Потім же він знову зв'язується і тим самим попереджає надлишкове зростання теломери.
З тієї ж причини теломерні ділянки ДНК малодоступні для інших ферментів - ДНК-метилаз і ендонуклеаз. У зв'язку з останньою обставиною, при мейозі в області теломер дуже низька частота дволанцюгових розривів.
Нарешті, за допомогою теломерних білків теломери кріпляться до компонентів ядерного матриксу в т. ч., можливо, до ядерної ламіни (пластинці, пов'язаній з внутрішньою ядерною мембраною). Чи дійсно в усіх клітинах теломери прикріплені до ядерної мембрани, поки не цілком ясно. Але, принаймні, на ранніх і середніх стадіях профази мейозу такий зв'язок, безперечно, існує.
Вважають також, що теломерна ДНК утворює декілька петель (у вигляді "пелюсток ромашки"), фіксованих на матриксі; і у міру укорочення теломер число "пелюсток" поступово зменшується.
Функції теломер
Тепер обговоримо відомі функції теломер.
1. Деякі функції можна умовно позначити як механічні.
а) Так, тільки що було сказано, що теломеры беруть участь у фіксації хромосом до ядерному матриксу. Це важливо для правильної орієнтації хромосом в ядрі, і ця обставина особливо проявляється в мейозі.
На зиготенній стадії профази мейозу відбуваються направленні переміщення кінців хромосом на поверхні ядерної мембрани - так, що кінці гомологічних хромосом змикаються і з них починається спаровування (кон'югація) цих хромосом строго однорідними ділянками.
б) Крім того, теломери зчіплюють один з одним кінці сестринських хроматид (що утворюються в хромосомі після S -фази). Можливо, це зчеплення відбувається за рахунок гібридизації теломер сестринської ДНК.
В той же час структура теломер така, що допускає розходження хроматид в анафазі. Проте можлива мутація (на рівні гена теломеразної РНК; див нижче), яка змінює нуклеотидну послідовність теломер; тоді розбіжність хроматид блокується.
2. Функції другої групи - стабілізаційні.
а) Найважливіша з них нам вже знайома: якщо в клітині немає теломерази (чи ALT), то наявність теломер оберігає від недореплікації генетично значущі відділи ДНК.
б) Якщо ж в клітині є теломеразна активність, то з’являється ще одна можливість стабілізації кінців разірванних хромосом.
Так, при випадковому розриві хромосоми утворюються фрагменти, на одному або на обох кінцях яких немає теломерних повторів. У відсутність теломерази ці фрагменти зливаються і деградують, що блокує клітинний цикл і приводить клітину до загибелі.
У присутності ж теломерази до місць розриву приєднується теломерна ДНК. Це стабілізує хромосомні фрагменти і дозволяє їм функціонувати.
Зокрема, цей феномен виявлений у хворих α-талассемиєю: в генах α-глобіну відбуваються розриви хромосоми 16q, і до пошкодженого кінця додаються теломерні повтори.
3.Вплив на експресію генів.
Ще одна цікава властивість теломер позначається як ефект положення: активність генів, розташованих поряд з теломерами, понижена (репресована). Такий ефект часто позначається як мовчання транскрипції, або сайленсинг.
При значному ж укороченні теломер ефект положення пропадає і прителомерні гени активуються.
а) Сайленсинг може бути результатом дії білків (таких, як Rapl або TFR1), що взаємодіють з теломерами. Тим більше, як вже відзначалося, ці білки знижують доступність теломерної ДНК для цілого ряду ферментів.
б) З іншого боку, ефект положення може бути обумовлений близькістю до ядерної оболонки. Так, по гіпотезі А. М. Оловнікова, в цій оболонці можуть розташовуватися Са2+ -канали, і потік іонів Са2+ впливає на взаємодію білків з довколишніми генами.
Ефект положення може торкнутися і внутрішніх генів, якщо який-небудь з таких генів стає транспозоном (геном, здатним до переміщення в іншу ділянку ДНК) і вбудовується в теломерну область. Чи якщо відбувається розрив хромосоми і утворення на кінцях розриву теломерних повторів : за допомогою останніх стає можливо зв’язeвання теломерних білків і прикріплення до ядерної мембрани.
4. "Рахункова" функція.
Теломерні відділи ДНК виступають годинниковим пристроєм (т. з. реплікометра), який відраховує кількість поділів клітини після зникнення теломеразної активності. Дійсно, як вже відзначалося, кожний поділ призводить до укорочення теломери на 50-65 н. п.
Причому набагато важливіше для клітини не те, скільки поділів вже пройшло, а скільки ще залишилося до критичного укорочення теломери. Тому можна сказати і так, що теломерний пристрій визначає кількість поділів, які здатна зробити нормальна клітина у відсутність теломерази.
Досягаючи ж критично короткої довжини, теломери втрачають можливість виконувати усі або більшість вищеперерахованих функцій. Порушується клітинний цикл, і кінець кінцем клітина гине.
Механізм дії теломерази
Із сказаного витікає дуже важлива біологічна роль теломеразы.
Теломераза подовжує G-ланцюг кожної теломери. Розберемося в механізмі цього процесу.
Ключову роль грає той факт, що з теломеразою пов'язана т. з. теломеразна РНК завдовжки близько 450 нуклеотидів. Її середня коротка ділянка комплементарна півтори теломерним повторам:
(3´)................АУЦ ЦЦА АУЦ...................(5´).
Лівий триплет цієї РНК (АУЦ) використовується для зв’язування (шляхом гібридизації) з крайнім теломерним напівповтором G-ланцюга ДНК (рис .1.19).
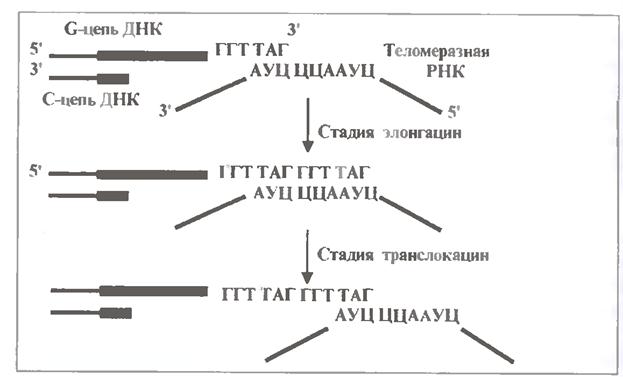
Інший же гексануклеотид (ЦЦААУЦ) служить в якості матриці для подовження 3'-кінця β-ланцюга на один теломерний повтор. А саме послідовне включення нуклеотидів (відповідно до структури матриці) каталізується білковими субодиницями теломерази.
Отже, теломераза виступає як зворотна транскриптаза - фермент, що здійснює синтез ДНК на РНК-матриці. В той же час напрям подовження ланцюга ДНК залишається звичайним: 5´ → 3'.
Процес утворення нового теломерного повтору позначають як елонгацію.
Потім відбувається транслокація - переміщення ферменту разом зі своєю РНК по подовжуваному ланцюгу ДНК на один повтор "лівіше", тобто до 3'-концу. Очевидно, в процесі транслокації РНК втрачає зв'язок з ДНК, а потім її встановлює, взаємодіючи з крайньою "половинкою" тільки що синтезованого повтору.
Потім ланцюг ДНК подовжується ще на один повтор. І так далі.
Таким чином, цикл роботи теломерази включає дві стадії - елонгацію і транслокацію. Чергуючи їх, фермент прибудовує до старого ланцюга декілька десятків або сотень теломерних повторів.
При цьому виявляється, що теломерна РНК - не лише матриця, але і важлива частина каталітичної машини : при заміні в ній декількох нуклеотидів відбувається не синтез "неправильних" повторів, а втрата теломеразою своєї активності.
Методи визначення активності теломерази
Визначають наявність теломерази в тому або іншому біологічному об'єктіза допомогою двох основних методів.
У т. з. прямому методі в інкубаційну суміш, окрім дослдіжуваного зразка, вносять праймер G -ланцюга (олігонуклеотид, зщо тримає декілька теломерных повторів) і мічені дНТФ. Праймер виступає в ролі G -цепи, з якою зв'язується і доторую потім подовжує теломераза. Про наявність активності судять по включенню радіоактивності до складу олігонуклеотида.
Другий, непрямий, метод - т. з. TRAP-аналіз (Telomer Repeat Amplification Protocol); він використовує ПЦР - полимераз-ную ланцюгову реакцію. Тут в суміш, окрім зразка, теж добавляют G -праймер і мічені нуклеотиди. Але, крім того, до суміші вносять С-праймер і термостійку бактерійну ДНК-полимеразу. Завдяки цьому, в ході ПЦР відбувається багатократне "клонування" (чи ампліфікація) подовженого тіломеразой олігонуклеотида.

Принцип ПЦР показаний на мал. 1.20.
З подовженою С-цепью взаємодіє С-праймер, після чого він нарощується ДНК-полимеразой (использующей С-цепь як матрицы). Утворюється двухце-почечная ДНК. Її ланцюги разівделяют нагріванням.
З подовженою С-цепью взаємодіє С-праймер, який потім нарощується ДНК-полимеразой. Утворюющуюся дволанцюжковий ланцюг знову розділяють нагреваниїм - в середовищі з'являється дополнительная подовжена С-цепь. Але тепер вона синтезирована вже не теломеразой, а ДНК-полимеразой - шляхом двустадийного копіювання першої С-цепи.
І так далі. Многократале чергуються цикли, вклющо сподіваються взаємодії С-или С-праймера з соответствующей поодиноким ланцюгом, полімеразного копіювання цього ланцюга і розподілу двох
Усе це відбувається в одній пробірці, оскільки термостабильная ДНК-полимераза витримує нагрівання у кінці кожного з численних циклів.
У результаті кількість С-цепей амплифицируется (множитьця) приблизно в 10' разів, і, відповідно, в стільки ж раз повышается чутливість методу.
Продукти реакції піддають гель-электрофорезу і підлозісподіваються спектру радіоактивних смуг, відповідних подовженийным олигонуклеотидам.
З появою ТКАР-аналізу різко полегшилося виявлення теломеразы в різних об'єктах. Проте і досі, в основному, йдеться лише про якісне виявлення активності. Стандартного ж способу виміру рівня активності до последнего часу не існувало.
Крім того, оцінюючи ті або інші результати, потрібно мати на увазі можливість артефактів. Одні обставини методического характеру можуть маскувати теломеразную активність; інші, навпаки, - давати псевдопозитивні результати.
Читайте також:
- III. Географічна структура світового ринку позичкового капіталу
- VІ. План та організаційна структура заняття
- Адміністративно – територіальний устрій і соціальна структура Слобожанщини у половині XVII – кінці XVIII століття
- Акти з охорони праці, що діють в організації, їх склад і структура.
- АРХІВНІ ДОВІДНИКИ В СИСТЕМІ НДА: ФУНКЦІЇ ТА СТРУКТУРА
- Атомно-кристалічна структура металів
- Базова алгоритмічна структура
- Банківська система та її структура. Функції Центрального банку.
- Безцехова виробнича структура.
- Будова систем: підсистема, елемент, структура, зв'язок.
- Бухгалтерська оцінка капіталу банку. Структура капіталу
- Бюджетна структура
| <== попередня сторінка | | | наступна сторінка ==> |
| Подовження теломер за допомогою теломерази | | |
|
Не знайшли потрібну інформацію? Скористайтесь пошуком google: |
© studopedia.com.ua При використанні або копіюванні матеріалів пряме посилання на сайт обов'язкове. |