РЕЗОЛЮЦІЯ: Громадського обговорення навчальної програми статевого виховання
ЧОМУ ФОНД ОЛЕНИ ПІНЧУК І МОЗ УКРАЇНИ ПРОПАГУЮТЬ "СЕКСУАЛЬНІ УРОКИ"
ЕКЗИСТЕНЦІЙНО-ПСИХОЛОГІЧНІ ОСНОВИ ПОРУШЕННЯ СТАТЕВОЇ ІДЕНТИЧНОСТІ ПІДЛІТКІВ
Батьківський, громадянський рух в Україні закликає МОН зупинити тотальну сексуалізацію дітей і підлітків
Відкрите звернення Міністру освіти й науки України - Гриневич Лілії Михайлівні
Представництво українського жіноцтва в ООН: низький рівень культури спілкування в соціальних мережах
Гендерна антидискримінаційна експертиза може зробити нас моральними рабами
ЛІВИЙ МАРКСИЗМ У НОВИХ ПІДРУЧНИКАХ ДЛЯ ШКОЛЯРІВ
ВІДКРИТА ЗАЯВА на підтримку позиції Ганни Турчинової та права кожної людини на свободу думки, світогляду та вираження поглядів
- Гідрологія і Гідрометрія
- Господарське право
- Економіка будівництва
- Економіка природокористування
- Економічна теорія
- Земельне право
- Історія України
- Кримінально виконавче право
- Медична радіологія
- Методи аналізу
- Міжнародне приватне право
- Міжнародний маркетинг
- Основи екології
- Предмет Політологія
- Соціальне страхування
- Технічні засоби організації дорожнього руху
- Товарознавство продовольчих товарів
Тлумачний словник
Авто
Автоматизація
Архітектура
Астрономія
Аудит
Біологія
Будівництво
Бухгалтерія
Винахідництво
Виробництво
Військова справа
Генетика
Географія
Геологія
Господарство
Держава
Дім
Екологія
Економетрика
Економіка
Електроніка
Журналістика та ЗМІ
Зв'язок
Іноземні мови
Інформатика
Історія
Комп'ютери
Креслення
Кулінарія
Культура
Лексикологія
Література
Логіка
Маркетинг
Математика
Машинобудування
Медицина
Менеджмент
Метали і Зварювання
Механіка
Мистецтво
Музика
Населення
Освіта
Охорона безпеки життя
Охорона Праці
Педагогіка
Політика
Право
Програмування
Промисловість
Психологія
Радіо
Регилия
Соціологія
Спорт
Стандартизація
Технології
Торгівля
Туризм
Фізика
Фізіологія
Філософія
Фінанси
Хімія
Юриспунденкция
Вікові особливості зовнішніх проявів діяльності серця.
Серцевий поштовх добре видний на око в дітей і підлітків з погано розвитою підшкірною жировою клітковиною, а в дітей з гарною угодованістю серцевий поштовх легко визначається при пальпації.
У немовлят і в дітей до 2–3літнього віку серцевий поштовх відчувається в 4-му лівому міжребер`ї на 1–2 см зовні від соскової лінії, у дітей 3–7літнього віку і наступні вікові групи він визначається в 5-му міжребер`ї, декілька варіюючи зовні й усередині від соскової лінії.
Тони серця в дітей декілька коротші в порівнянні з дорослими. Якщо в дорослих перший тон триває 0,1–0,17 сек., то в дітей 0,1–0,12 сек.
Другий тон у дітей більш тривалий, чим у дорослих. У дітей він триває 0,07–0,1 сек., а в дорослих – 0,06– 0,08 сек. Іноді в дітей від 1 до 3 років спостерігається розщеплення другого тону, зв’язане з дещо різночасним закриттям напівмісячних клапанів аорти і легеневої артерій, і розщеплення першого тону, що обумовлено асинхронним закриттям митрального і тристулкового клапанів.
Нерідко в дітей реєструється третій тон – дуже тихий, глухий і низький. Він виникає на початку діастоли через 0,1–0,2 сек. після другого тону і зв’язаний зі швидким розтяганням м’яза шлуночків, що виникає при надходженні в них крові. У дорослих третій тон триває 0,04–0,09 сек., у дітей – 0,03–0,06 сек. У новонароджених і грудних дітей третій тон не вислуховується.
Під час м’язової роботи, позитивних і негативних емоцій збільшується сила серцевих тонів, під час сну вона зменшується.
Електрокардіограма дітей значно відрізняється від електрокардіограми дорослих і в різні вікові періоди має свої особливості в зв’язку зі зміною розмірів серця, його положення, регуляції й ін.
У плоду електрокардіограма реєструється на 15–17-й тиждень вагітності.
Час проведення збудження від передсердь у шлуночки (інтервал Р-Q) у плоду більш короткий, ніж у немовляти. У немовлят і дітей перших трьох місяців життя цей час дорівнює 0,09–0,12 сек., а в більш старших дітей – 0,13–0,14 сек.
Комплекс QRS у новонароджених коротший, ніж у більш старшому віці. Окремі зубці електрокардіограми в дітей цього віку різні в різних відведеннях.
У грудних дітей в електрокардіограмі залишається сильно вираженим зубець Р, що пояснюють більшою величиною передсердь. Комплекс QRS часто багатофазний, у ньому переважає зубець R. Зміни комплексу QRS зв’язують з нерівномірним ростом провідної системи серця.
У дошкільному віці електрокардіограма більшості дітей цього віку характеризується деяким зменшенням зубців Р и Q. Зубець R збільшується у всіх відведеннях, що зв’язано з розвитком міокарда лівого шлуночка. У цьому віці збільшується тривалість комплексу QRS і інтервалу Р – Q, що залежить від закріплення впливів блукаючого нерва на серце.
У дітей шкільного віку ще більше збільшується тривалість серцевого циклу (R – R) і в середньому дорівнює 0,6–0,85 сек. Величина зубця R у першому відведенні в підлітків наближається до величини його у дорослого. Зубець Q зменшується з віком, і в підлітків також наближається до величини його в дорослого.
У шкільному, віці ще більше збільшується тривалість комплексу QRS, що відбиває вікове зменшення частоти серцевих скорочень.
ВІКОВІ ОСОБЛИВОСТІ СИСТОЛІЧНОГО
І ХВИЛИННОГО ОБ`ЄМІВ СЕРЦЯ
Величина систолічного об`єму серця з віком збільшується більш значно, чим величина хвилинного об`єму. На зміні хвилинного об`єму позначається зменшення числа серцевих скорочень з віком.
Величина хвилинного об`єму в немовляти і дітей до 1 року в середньому дорівнює 0,33 л, у віці 1 року – 1,2 л, у 5-річних дітей – 1,8 л, у 10-річних – 2,5 л. Величина систолічного об`єму в немовляти дорівнює 2,5 мл, а в дітей 1 року – 10,2 мл.
Хвилинний і систолічний об`єми серця залежать не тільки від віку, але і зв’язані з загальним фізичним розвитком дітей. У дітей, більш розвитих фізично, більша величина систолічного і хвилинного об`ємів.
У дівчаток і жінок величина хвилинного і систолічного об`ємів менша, ніж у юнаків і чоловіків.
Хвилинний об`єм серця зв’язаний з потребами організму в кисні. До кінця першого року життя кількість спожитого кисню на 1 кг маси в дітей в 2–3 рази більше, ніж у дорослого.
Відповідно, і хвилинний об`єм крові на 1 кг маси дітей цього віку більше в 2 із зайвим разу, чим у дорослих.
При виконанні різної фізичної роботи хвилинний об`єм серця в дітей збільшується в меншому ступені, чим у дорослих.
У дітей усіх вікових груп збільшення хвилинного об`єму при помірному фізичному навантаженні відбувається за рахунок збільшення систолічного обсягу. При навантаженнях максимальної потужності мобілізуються всі сили організму і хвилинний об`єм збільшується за рахунок збільшення як систолічного об`єму, так і числа серцевих скорочень.
ВІКОВІ ОСОБЛИВОСТІ РУХУ КРОВІ ПО СУДИНАХ
Особливості змін кров’яного тиску з віком. У новонародженої дитини середня величина кров’яного тиску складає 76 мм рт. cm. У дітей усіх віків мається загальна тенденція до збільшення систолічного, діастолічного і пульсового тиску з віком.
Максимальний кров’яний тиск до 1 року дорівнює 100 мм рт. ст., до 5–8 років – 104 мм рт. ст., до 11–13 років – 127 мм рт. ст., до 15–16 років – 134 мм рт. ст. Мінімальний тиск, відповідно, дорівнює: 49, 68, 83 і 88 мм рт. cm. (за даними А. М. Попова).
Величина артеріального тиску в дітей одного віку значно коливається. Більш високий тиск відзначений у дітей, що мають більший ріст і масу.
Величина кров’яного тиску в дітей легко змінюється під впливом різних зовнішніх факторів. Так, при переході тіла з положення сидячи в горизонтальний кров’яний тиск у більшості дітей підвищується на 10–20 мм рт. ст.
На величину кров’яного тиску в дітей впливають кліматичні і географічні умови місцевості: у дітей усіх вікових груп, що проживають на півдні, кров’яний тиск нижче, ніж у жителів півночі.
Кров’яний тиск у дітей різко змінюється під впливом емоцій: максимальний тиск підвищується на 20–40 мм рт. ст., мінімальне – на декілька меншу величину.
У грудних дітей відзначене підвищення тиску при прийомі їжі. Ранком кров’яний тиск нижче, а до вечора підвищується.
Заняття в школі впливають на величину кров’яного тиску учнів. На початку навчального дня відзначене підвищення мінімального тиску від уроку до уроку і зниження максимального (тобто зменшення пульсового тиску). До кінця навчального дня кров’яний тиск підвищується. При наявності уроків праці і фізкультури відзначене менше зниження величини пульсового тиску.
Під час м’язової роботи в дітей підвищується величина максимального і декілька знижується величина мінімального тиску. Під час виконання граничного м’язового навантаження в підлітків і юнаків величина максимального кров’яного тиску може зростати до 180–200 мм рт. ст. Оскільки в цей час величина мінімального тиску змінюється незначно, то й пульсовий тиск зростає до 50–80 мм рт. ст., що говорить про збільшення сили скорочення серця. Інтенсивність змін величини кров’яного тиску під час фізичного навантаження залежить від віку: чим старша дитина, тим значніше ці зміни.
Вікові зміни кров’яного тиску при фізичному навантаженні особливо яскраво виявляються у відновлювальному періоді. Відновлення величини систолічного тиску до вихідної величини здійснюється тим швидше, ніж старше вік дітей.
Пульсовий тиск у дітей коливається у великих межах.
Величина венозного тиску зменшується з віком. Якщо в дітей перших років життя воно дорівнює 105 мм вод. ст., то в підлітка воно знижується до 86 мм вод. cm. Його величина коливається у великих межах.
Велика величина венозного тиску в маленьких дітей зв’язана з великою кількістю циркулюючої в організмі крові, вузьким просвітом вен і зниженою їхньою ємністю. Вона залежить від сили скорочень правого шлуночка і судинного тонусу. Венозний тиск у дітей не залежить від частоти серцевих скорочень і від коливань максимального і мінімального артеріального тиску.
Величина венозного тиску зв’язана з фазами подиху: при вдиху воно декілька знижується, а при видиху – підвищується.
Воно різко збільшується під час негативних емоцій. Наприклад, при плачі венозний тиск у дітей може підвищитися до 335мм вод. Ст.
Вікові зміни швидкості руху крові. З віком швидкість руху крові сповільнюється. У немовлят кров робить кругообіг за 12 сек., у 3-річних – за 15 сек., у дітей 7-8 років за 7–8 сек., у 14-річних – за 18 ½ сек. Уповільнення швидкості руху крові зв’язано з віковими змінами судин, перед усе зі збільшенням їх довжини в зв’язку з ростом дитини. На швидкість руху крові впливає і зміна частоти серцевих скорочень: зменшення числа серцевих скорочень з віком приводить до уповільнення швидкості руху крові.
У всі вікові періоди в жінок швидкість руху крові по судинах більша, ніж у чоловіків.
ВІКОВІ ОСОБЛИВОСТІ РЕГУЛЯЦІЇ
ДІЯЛЬНОСТІ СЕРЦЕВО-СУДИННОЇ СИСТЕМИ
Особливості регуляції в антенатальному періоді. Регуляція діяльності серця здійснюється в плоду симпатичною нервовою системою. Насамперед розвивається та частина симпатичної нервової системи, що регулює діяльність серця. Причому це відбувається задовго до того, як інші органи одержують симпатичну іннервацію. Насамперед розвиваються клітини в зірчастому ганглії.
Центр симпатичної регуляції серця знаходиться в постійній тонічній напрузі, що залежить головним чином від змісту кисню в крові плоду. Ступінь насичення киснем крові плоду близько 50%, тобто кров плоду є в значній мірі гіпоксемічною у порівнянні з кров’ю немовляти. Така кров, що омиває клітини центра симпатичної регуляції серця, створює в них постійне тонічне збудження. Ці впливи обумовлюють велику частоту серцевих скорочень у плода.
Вікові особливості регуляції серцево-судинної системи в постнатальному періоді. У новонародженої дитини цілком сформовані еферентні нерви, що регулюють діяльність серця. У нього добре розвинуті розгалуження і периферичні закінчення блукаючого і симпатичного нервів. Але проте і в цьому віці серце регулюється в основному тільки симпатичними нервами.
Після народження на ранніх етапах постнатального розвитку залишається превалюючим тонічний вплив центра симпатичної регуляції серця. Але в цей час тонічна напруга центра підтримується не гуморальним шляхом, а рефлекторно-аферентною проприоцептивною імпульсацією. Причому м’язовий тонус залежить від температури навколишнього середовища. Він найкращий при температурі 20° С. При підвищенні температури навколишнього середовища знижується тонус скелетної мускулатури і при досягненні 28–30°С м’яза розслаблюються; слідом за цим знижується тонус центра симпатичної іннервації серця і зменшується частота серцевих скорочень. При зменшенні температури навколишнього середовища підвищується м’язовий тонус і тонус центра симпатичної регуляції серця, внаслідок чого збільшується частота серцевих скорочень.
Відсутність парасимпатичної регуляції діяльності серця зв’язують з недостатньою розвиненістю центра блукаючого нерва.
Посилення впливів блукаючого нерва на серце пояснюють установленням постійного тонічного збудження його центра. І.А.Аршавский вважає, що в цьому процесі велике значення має розвиток скелетної мускулатури. Показано, що перші ознаки впливів блукаючого нерва спостерігаються в той період, коли дитина починає утримувати голівку, що має місце в 2,5– 3-місячному віці. У 3–4-річному віці збільшується рухова активність дітей, і в цей період відзначається закріплення впливів блукаючого нерва на серце.
У період встановлення впливів блукаючого нерва на серце зменшується ступінь поляризації синусного вузла, збільшується тривалість одного циклу серцевих скорочень, внаслідок чого зменшується їх число за одиницю часу. У цей період характер електрокардіограми і частота серцевих скорочень стають майже такими ж, як у дорослих.
Відзначено, що в періоди появи і підвищення тонусу центра блукаючого нерва підвищується тонічне збудження судинорухального центру.
РОЗДІЛ ХІІІ ДИХАЛЬНА СИСТЕМА
ЗНАЧЕННЯ ДИХАННЯ. БУДОВА ОРГАНІВ ДИХАННЯ
Значення дихання, його основні етапи. Дихання забезпечує безупинне постачання всіх органів і тканин тіла киснем і видалення з організму вуглекислого газу, що постійно утворюється в процесі обміну речовин.
Органи дихання мають захисну функцію. Повітроносні шляхи вистилає слизова оболонка, що містить велику кількість окремих клітин і залоз, що виділяють слиз, що зволожує поверхню повітроносних шляхів. Цей слиз володіє і бактерицидними властивостями – у ньому міститься лізоцим – речовина, що знижує здатність бактерій до розмноження чи убиває їх.
На поверхню слизової виходять із кровоносних судин лейкоцити, що теж виконують захисну функцію. Здійснюючи фагоцитоз, вони гинуть, і тому в слизу, що виділяється з носа, міститься багато загиблих лейкоцитів. До слизу, що покриває повітроносні шляхи, прилипають частки пилу, що містяться в атмосферному повітрі. Затримці сторонніх часток сприяють і волоски, що знаходяться біля зовнішнього краю носових отворів.
Більшість клітин слизової оболонки вкриті численними рухливими війками. Вони постійно хвилеподібно коливаються, причому в зовнішньому напрямку швидше, ніж у внутрішньому. Це забезпечує проштовхування слизу і різних механічних часток назовні.
Повітроносні шляхи носової порожнини мають велику мережу капілярів, завдяки чому повітря зігрівається теплом, що віддається кров’ю. Таким чином, у легені попадає зігріте і чисте повітря, звільнене від бактерій і сторонніх механічних часток.
У процесі дихання розрізняють декілька етапів: 1) обмін газів між органами дихання і зовнішнім середовищем; 2) обмін газів у легенях (між альвеолярним повітрям і кров’ю легеневих капілярів);
3) перенос газів кров’ю; 4) обмін газів у тканинах (між тканинами і артеріальною кров’ю, що притікає по них); 5) клітинне дихання (споживання клітинами кисню і виділення вуглекислоти).
Етап дихання, що полягає в обміні газів між зовнішнім середовищем і органами дихання, одержав назву зовнішнього дихання. Крім зовнішнього дихання, розрізняють і внутрішнє дихання – обмін газів між тканинами і кров’ю.
Загальний план будови органів дихання. Органи дихання людини представлені повітроносними шляхами, по яких проходить вдихуване і видихуване повітря, і легенями, де відбувається обмін газів.
Дихальні шляхи починаються носовою порожниною, що відділена від ротової порожнини попереду твердим, а позаду м’яким піднебінням. Носова порожнина має кістковий і хрящовий кістяк і суцільною пертинкою поділяється на праву і ліву частини. Носові раковини розділяють носову порожнину на ряд вузьких носових щілин, по яких проходить вдихуване і видихуване повітря.
З носової порожнини повітря проходить у носоглотку, звідки воно переходить у носову частину глотки, а потім у гортань.
Гортань розташовується попереду гортанної частини глотки на рівні IV-VI шийних хребців і утворена хрящами: щитовидним, двома черпаловидними і перснюватий. До верхнього краю щитовидного хряща прикріплюється надгорляник, що закриває вхід у гортань під час ковтання і тим перешкоджає потраплянню в неї їжі. Від щитовидного хряща до черпаловидного (попереду назад) йдуть дві голосові зв’язки. Простір між ними називають голосовою щілиною.
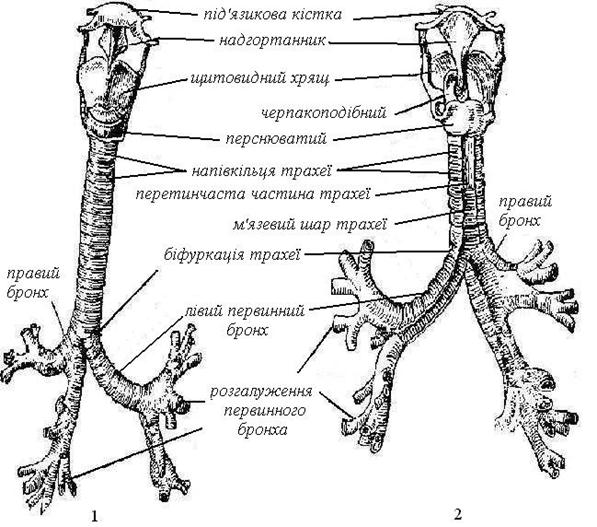
Рис. 88. Гортань, трахея і великі бронхи попереду (1) і позаду (2).
На рівні VI–VII шийних хребців гортань переходить у трахею, що має середню довжину 12 см. Трахея поділяється на два бронхи, що входять у праву і ліву легені. Трахея і бронхи складаються з хрящових півкілець, не замкнутих на задньій поверхні. Хрящові півкільця додають пружність дихальним шляхам і роблять їх незпадаючимися і тим самим легко прохідними для повітря.
У правій і лівій легені бронхи деревоподібно розгалужуються на більш дрібні бронхи, що входять у легеневі часточки й утворюють ще більш дрібні дихальні розгалуження – бронхіоли. Дрібні дихальні бронхіоли діаметром близько 0,5 мм розгалужуються на альвеолярні ходи, що закінчуються альвеолярними мішечками. Альвеолярні ходи і мішечки на стінках мають випинання у вигляді пухирців, що називають альвеолами. Діаметр альвеол дорівнює 0,2–0,3 мм, а їх кількість досягає 300-400 млн., завдяки чому створюється велика дихальна поверхня легень. Вона досягає 100-120 м2.
Альвеоли складаються з дуже тонкого плоского епітелію, що зовні оточений мережею дрібних, теж тонкостінних, кровоносних судин, що полегшує обмін газів.
Розташування легень у грудній порожнині. Для розуміння механізму вдиху і видиху важливо добре представити положення легень у грудній порожнині. Легені розташовуються в герметично закритій грудній порожнині. Задня стінка грудної порожнини утворена грудним відділом хребта і рухливо приєднаними до хребців ребрами. З боків вона утворена ребрами, попереду – ребрами і грудиною. Між ребрами розташовуються міжреберні м’язи (зовнішні і внутрішні). Знизу грудна порожнина відокремлюється від черевної діафрагмою, куполоподібно вигнутою в грудну порожнину.
У людини дві легені – права і ліва. Права легеня складається з трьох часток, ліва – із двох. Звужену верхню частину легень називають верхівкою. Розрізняють ворота легені – поглиблення на їх внутрішній поверхні, через яке проходять бронхи, кровоносні судини (легенева артерія і дві легеневі вени), лімфатичні судини і нерви. Сукупність цих утворень зветься коренем легені.
Легені покриті оболонкою – плеврою, що складається з двох листків: внутрішнього (вісцерального) і зовнішнього (парієтального). Внутрішній листок плеври вкриває легені і є їх зовнішньою оболонкою, яка по кореню легко переходить у зовнішній листок плеври, що вистилає стінки грудної порожнини (є її внутрішньою оболонкою). Таким чином, між внутрішнім і зовнішнім листками плеври утворюється герметично замкнений дрібний капілярний простір, що називають плевральною порожниною. У ній знаходиться невелика кількість (1–2 мл) плевральної рідини, що змочує листки плеври і полегшує їх ковзання відносно один одного.
Однією з основних причин зміни повітря в легенях є зміна об’єму грудної і плевральної порожнин. Легені пасивно рухаються за зміною їх об’єму.
ЗОВНІШНЄ ДИХАННЯ
Механізм акта вдиху і видиху. Акт вдиху (інспірація) – активний процес, у здійсненні якого важливе значення має зміна об’єму грудної порожнини.
Оскільки легені постійно з’єднані через повітроносні шляхи із зовнішнім середовищем, то тиск повітря у них дорівнює атмосферному. Легені знаходяться завжди в розтягнутому стані. Під час вдиху їх розтягнення збільшується, під час видиху – зменшується.
У здійсненні вдиху беруть участь зовнішні міжреберні м’язи і діафрагма. При скороченні зовнішніх міжреберних м’язів, що йдуть зверху вперед і вниз, ребра піднімаються, і при цьому збільшується об’єм грудної порожнини.
Діафрагма, скорочуючись, займає більш плоске положення, завдяки чому об’єм грудної порожнини збільшується. Якщо збільшується об’єм герметично закритої порожнини, то тиск у ній буде зменшуватися, що і відбувається в грудній порожнині. Внаслідок зменшення тиску в грудній порожнині створюється еластична тяга легень: легені розширюються і у них спрямовується повітря – відбувається вдих.
Здійсненню вдиху сприяє зменшення тиску в плевральній порожнині. При спокійному дихання тиск у плевральній порожнині дорівнює мінус 6-8 мм рт. ст., тобто на 6-8 мм рт. ст. нижче атмосферного. Його можна вимірити, якщо ввести в щілину голку, з’єднану з манометром.
У момент вдиху, коли скорочуються зовнішні міжреберні м’язи і ребра піднімаються, зовнішній листок плеври відходить від внутрішнього, унаслідок чого збільшується об’єм плевральної порожнини, а тиск у ній зменшується. Чим глибше вдих, тим більше зменшується тиск. У момент глибокого вдиху воно може досягати мінус 30 мм. рт. ст. Зменшення тиску в плевральній порожнині приводить до того, що внутрішній листок плеври спрямовується до зовнішнього, внаслідок чого легені розширюються й у них засмоктується повітря.
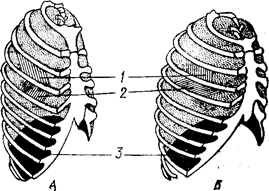
|
Рис. 88.А. Грудна клітка при видиху (А) і вдиху (5):
1 – зовнішні міжреберні м’язи;
2 – внутрішні міжреберні м’язи;
3 – діафрагма.
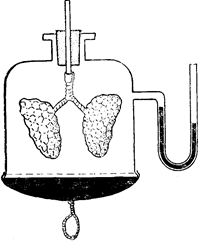
|
Рис. 88. Б. Модель Дондерса для демонстрації механізму вдиху і видиху.
У різних людей переважне значення в здійсненні акта вдиху можуть мати міжреберні м’язи чи діафрагма. Тому говорять про різні типи дихання: грудне (чи реберне) і черевне (чи диафрагмальне). Тип дихання не є яскраво вираженим, частіше має місце змішаний дихання, і його тип виявляється в залежності від умов. Так, під час вагітності в жінок утруднене діафрагмальне дихання і переважає грудне.
Акт видиху (експірація) – пасивний процес. Коли в міжреберних м’язах закінчується процес збудження, вони розслаблюються, унаслідок чого ребра пасивно повертаються у вихідне положення, точно так само припинення скорочення діафрагми приводить до того, що вона займає своє колишнє куполообразное положення. Повернення ребер і діафрагми у вихідне положення приводять до зменшення об’єму грудної порожнини, а отже, до зменшення в ній тиску. Одночасно при поверненні ребер у вихідне положення тиск у плевральній порожнині підвищується, тобто в ній зменшується негативний тиск. При глибокому видиху воно дорівнює –3, –4 мм рт. ст. Усі ці процеси, що забезпечують підвищення тиску в грудній і плевральній порожнині, приводять до того, що легені здавлюються і з них пасивно виходить повітря – здійснюється видих.
Посилений видих є активним процесом. У його здійсненні беруть участь: внутрішні міжреберні м’язи, волокна які йдуть у протилежному напрямку в порівнянні з зовнішніми: знизу нагору і вперед. При їх скороченні ребра опускаються вниз і об’єм грудної порожнини зменшується. Посиленому видиху сприяє також скорочення м’язів черевного преса, що давлять на діафрагму, збільшуючи її купол, зменшують об’єм грудної порожнини. Нарешті, м’язи пояса верхніх кінцівок, скорочуючись, здавлюють у верхній частині грудну клітку і зменшують її об’єм.
У результаті зменшення об’єму грудної порожнини в ній збільшується тиск, унаслідок чого повітря виштовхується з легень – відбувається активний видих. На вершині видиху тиск у легенях може бути більше атмосферного на 3-4 мм рт. ст.
Акти вдиху і видиху ритмічно змінюють один одного. Доросла людина робить 15-20 дихальних рухів за хвилину. Дихання фізично тренованих людей більш рідке (до 8-12 дихальних рухів за хвилину) і глибоке.
Життєва ємність легень. Життєвою ємністю легень називають ту кількість повітря, що може людина видихнути після максимального вдиху чи максимально вдихнути після максимального видиху.
Життєва ємність легень визначається приладом спірометром, а метод її визначення називають методом спірометрії. При визначенні життєвої ємності людина після максимального вдиху робить посилений видих через трубку в спірометр, по шкалі якого відраховують величину життєвої ємності легень.
Величину життєвої ємності легень складають декілька об’ємів повітря: дихальний (500 см3), додатковий (1500 см3) і резервний (1500 см3).
Дихальним об’ємом легень називається та кількість повітря, що людина видихає при спокійному диханні. Воно дорівнює в середньому 500 см3. Але після спокійного видиху людина може ще видихнути (не роблячи додаткового вдиху) якийсь певний об’єм повітря, що залишилося в легенях. Та кількість повітря, що людина може видихнути після спокійного видиху, називають резервним об’ємом повітря. Його величина в середньому складає 1500 см3.
Здійснивши спокійний вдих, людина може ще посилено вдихнути (не роблячи видиху після спокійного вдиху) близько 1500–2000 см3 повітря. Цю кількість повітря називають додатковим об’ємом. Його спірометром не визначають, а розраховують, віднімаючи з життєвої ємності суму дихального і резервного об’ємів.
Життєва ємність легень у середньому у дорослої людини дорівнює 3500 – 4000 см3, у чоловіків вона дещо більша, ніж у жінок.
Життєва ємність легень не характеризує всього об’єму повітря, що знаходиться в легенях. Після того як людина максимально видихає, у неї в легенях залишається велика кількість повітря. Воно складає 1000-1200 см3, і називають його залишковим об’ємом повітря. Цей об’єм разом з резервним об’ємом складає нормальну ємність легень.
З кожним актом дихання не весь дихальний об’єм повітря попадає в легені. Значна частина його (160-180 см3) залишається у повітроносних шляхах (у носоглотці, трахеї, бронхах). Об’єм повітря, що заповнює великі повітроносні шляхи, називають повітрям «мертвого» простору. У ньому не відбувається обмін газів. Таким чином, у легені з кожним вдихом попадає 500-160=340 см3 повітря. Цей об’єм приходить до наявного у легенях повітря, що складає нормальну ємність легень.
Виходить, що з кожним вдихом обмінюється: 340/2500=1/7 частина повітря.
Атмосферне повітря перед тим як потрапити в легені змішується з повітрям мертвого простору внаслідок чого вміст газів у ньому змінюється. По цій же причині неоднаковий вміст газів у видихуваному і альвеолярному повітрі.
Склад повітря (у %)
| Назва газу | Повітря | ||
| атмосферний | видихуваний | альвеолярний | |
| CO2 | 0,03 | 4,1 | 5,5-5,7 |
| O2 | 20,95 | 16,4 | 14,2-14,6 |
| Азот | 79,02 | 79,5 |
Легенева вентиляція. Безупинну зміну повітря, що відбувається в легенях, називають легеневою вентиляцією. Її показником може бути хвилинний об’єм легень, тобто кількість повітря, видихувана за хвилину. Величина хвилинного об’єму визначається добутком числа дихальних рухів у хвилину на об’єм одного вдиху (350-500 см3).
У жінок величина хвилинного об’єму може дорівнювати 3-5 л, а в чоловіків – 6-8 л. Його величина залежить від віку, статі і фізичного розвитку людини (його маси і зросту), рівня окислюальних процесів. Хвилинний об’єм значно збільшується при фізичній роботі і може досягати 50-100 л/хв. Величину вентиляції, що покриває потреби організму в стані спокою, називають ейпне (“ей” – добре, “пное” – дихання: “гарне дихання”).
Величина хвилинного об’єму легеневої вентиляції недостатньо відбиває ефективність дихання. При одній і тій же величині хвилинного об’єму в легенях може обмінюватися різна кількість повітря. Так, спортсмен за хвилину може здійснювати 10 глибоких дихань при об’ємі дихальних рухів 600 см3, тобто хвилинний об’єм у нього буде дорівнювати 6000 см3.
Нетренована людина може зробити 20 дихань, але поверхневих (дихальний об’єм дорівнює 300 см3) – величина хвилинного об’єму буде теж дорівнювати 6000 см3. Але у спортсмена в легені попадає 600 см3 – 160 см3 = 440 см3, тобто 3/4 дихального об’єму. У нетренованої людини обмінюється значно менше повітря: 300 см3 -160 см3 = 140 см3, тобто 1/2 дихального об’єму. З цього можна зробити висновок, що рідке, але глибоке дихання більш ефективне, тому що при цьому в альвеолярному повітрі здійснюється більший обмін газів.
ПЕРЕНЕСЕННЯ ГАЗІВ КРОВ’Ю
Значення фізичних факторів для переносу газів чкровыо. Розчинення газів у рідинах залежить від ряду факторів: від властивостей самого газу, від властивостей рідини (концентрації в ній солей,їїтемператури), від об’єму і тиску газу над рідиною.
Показником розчинності газів служить коефіцієнт розчинності (чи абсорбційний коефіцієнт). Його величина показує той об’єм газу, що розчиняється в 1 см3 рідині при температурі 0°С и тиску 760 мм рт. ст.
Коефіцієнт розчинності газу тим більше, чим нижче температура; він зменшується з підвищенням температури і при температурі кипіння дорівнює нулю (газ з розчину весь випаровується). Коефіцієнт розчинності в крові для кисню дорівнює 0,022, для азоту – 0,011, для вуглекислоти – 0,511.
У стані розчинення в артеріальній крові міститься 0,25 мл О2, 2,69 мл СО2 і 1,04 мл N.
Фізичне розчинення газів дуже мале, а тому воно не має великого значення для їхнього переносу кров’ю. Важливим фактором переносу газів кров’ю є утворення хімічних сполук з речовинами плазми крові і еритроцитів. Для встановлення хімічних зв’язків і фізичного розчинення газів важлива величина тиску газу над рідиною.
Роль тиску газів у їх переносі кров’ю. Надходженнягазув рідину залежить від його тиску. Якщо над рідиною знаходиться суміш газів, то рух і розчинення кожного з них залежатьвідйого парціального тиску. Парціальний тиск можна розрахувати виходячи із загального тиску суміші газів і їх процентного вмісту.
Усю газову суміш атмосферного повітря приймають за 100%, вона має тиск 760 мм рт. cm, а частина газу (O2 – 20,95%) приймають за х. Звідси: х = 760 · 20,95/100 = 159,22 мм рт. ст. При розрахунку парціального тиску газів в альвеолярному повітрі необхідно враховувати, що він насичений водяними парами, тиск яких складає 47 мм рт. ст. Отже, на частку газової суміші, що входить до складу альвеолярного повітря приходиться тиск не 760 мм рт. ст., а 760-47 = 713 мм рт. cm. Цей тиск приймається за 100%.
Звідси легко обчислити, що парціальний тиск О2, що міститься в альвеолярному повітрі в кількості 14,3%, буде рівний:
713 · 14,3/100=102 мм рт. ст.
Відповідний розрахунок парціального тиску СО2 показує, що воно дорівнює 40 мм рт. ст.
Альвеолярне повітря контактує з тонкими стінками легеневих капілярів, по яких приходить до легень венозна кров. Інтенсивність обміну газів і напрямок їхнього руху (з легень у кров чи із крові в легені) залежать від парціального тиску кисню і вуглекислоти в газовій суміші в легенях і в крові (тиск газів у рідинах називають їх напругою).
Напруга кисню у венозній крові дорівнює 40 мм. рт. ст., вуглекислоти – 46 мм рт. ст. Рух газів здійснюється від більшого тиску до меншого. Отже, кисень буде надходити з легень (його парціальний тиск у них дорівнює 102 мм рт. cm.) у кров (його напруга в крові 40 мм рт. ст.), а вуглекислий газ із крові (напруга 46 мм рт. ст.) в альвеолярне повітря (тиск 40 мм рт. ст.).
Киснева ємність крові. Вміст газів у крові. У крові кисень з’єднується з гемоглобіном і утворить неміцне з’єднання – оксигемоглобін. Насичення крові киснем залежить від кількості гемоглобіну в крові. Максимальна кількість кисню, що може поглинути 100 мл крові, називають кисневою ємністю крові. Відомо, що в 100 мл крові людини міститься 14% гемоглобіну. Кожен грам гемоглобіну може зв’язати 1,34 мл O2. Виходить, 100 мл крові можуть перенести 1,34 х 14% = 19 мл (19 об’ємних відсотків). Це і є киснева ємність крові.
Можна розрахувати ступінь насичення крові киснем. Для цього потрібно розділити вміст кисню досліджуваної крові на її кисневу ємність.
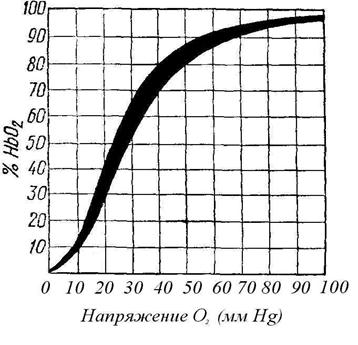
Зв’язок кисню з гемоглобіном. В артеріальній крові 0,25 об’ємного відсотка О2 знаходиться в стані фізичного розчинення в плазмі, а інші 18,75 об’ємного відсотка – в еритроцитах у пов’язаному стані з гемоглобіном у вигляді оксигемоглобіну. Зв’язок гемоглобіну з киснем залежить від величини напруги газів: якщо воно збільшується, гемоглобін приєднує кисень і утвориться оксигемоглобін (НВО2). При зменшенні напруги кисню оксигемоглобін розпадається і віддає кисень. Криву, що відбиває залежність насичення гемоглобіну киснем від напруги останнього, називають кривою дисоціації оксигемоглобіну. На малюнку видно, що навіть при невеликому парціальному тиску кисню (40 мм рт. ст.) з ним зв’язуються 75-80% гемоглобіну. При тиску 80 – 90 мм рт. cm. гемоглобін майже цілком насичується киснем. В альвеолярному повітрі парціальний тиск кисню дорівнює 120 мм рт. ст., тому кров у легенях буде цілком насичена киснем.
При розгляді кривої дисоціації оксигемоглобіну можна помітити, що при зменшенні парціального тиску кисню оксигемоглобін піддається дисоціації і віддає кисень. При нульовому тиску кисню оксигемоглобін може віддати весь з’єднаний з ним кисень.
Рис. 89 Залежність насичення крові людини киснем
від його парціального тиску (крива дисоціації оксигемоглобіну)
 |
| Рис. 90 Криві насичення гемоглобіну киснем при різних умовах: А – у залежності від реакції середовища (рН); Б – від температури; В – від вмісту солей; Г– від вмісту CO2. По осі абсцис – парціальний тиск О2; (у мм рт. ст.), по oсі ординат – ступінь насичення (у %). |
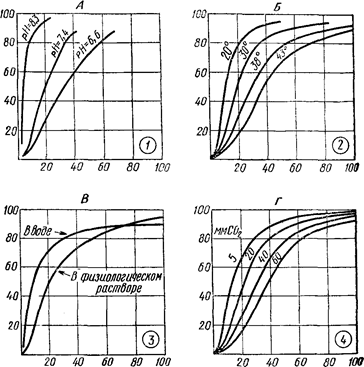
Властивість гемоглобіну – легко насичуватися киснем, навіть при невеликих тисках, і легко його віддавати – дуже важлива.
Завдяки легкій віддачі гемоглобіном кисню при зниженні парціального тиску забезпечується безперебійне постачання тканин киснем, у яких унаслідок постійного споживання кисню його парціальний тиск дорівнює нулю.
Розпад оксигемоглобіну на гемоглобін і кисень збільшується з підвищенням температури тіла.
Дисоціація оксигемоглобіну залежить від реакції середовища плазми крові. Зі збільшенням кислотності крові зростає дисоціація оксигемоглобіну.
Зв’язки гемоглобіну з киснем у воді здійснюється швидко, але повного його насичення не досягається, так само як не відбувається повної віддачі кисню при зниженні його парціального тиску. Більш повне насичення гемоглобіну киснем і повною його віддачею при зниженні напруги кисню відбуваються в розчинах солей і в плазмі крові.
Особливе значення в зв’язуванні гемоглобіну з киснем має вміст CO2 у крові. Чим більше міститься вуглекислоти в крові, тим менше зв’язується гемоглобін з киснем і тем швидше відбувається дисоціація оксигемоглобіну. Особливо різко знижується здатність гемоглобіну з’єднуватися з киснем при тиску СО2 рівному 46 мм рт. ст., тобто при величині, що відповідає напрузі СО2 у венозній крові. Вплив CO2 на дисоціацію оксигемоглобіну дуже важливий для переносу газів у легенях і тканинах.
У тканинах міститься велика кількість CO2 і інших кислих продуктів розпаду, що утворяться в результаті обміну речовин. Переходячи в артеріальну кров тканьових капілярів, вони сприяють більш швидкому розпаду оксигемоглобіну і віддачі кисню тканинам.
У легень же, у міру виділення СО2 з венозної крові в альвеолярне повітря, зі зменшенням вмісту CO2 у крові збільшується здатність гемоглобіну з’єднуватися з киснем. Тим самим забезпечується перетворення венозної крові в артеріальну.
Зв’язки вуглекислого газу кров’ю. В артеріальній крові міститься 50-52% СО2, а у венозній на 5-6% більше – 55-58%. З них 2,5-2,7 об’ємні відсотки в стані фізичного розчинення, а інша частина СО2 переноситься у вигляді солей вугільної кислоти: бікарбонату натрію (NaHСО3) у плазмі і бікарбонату калію (КНСО3) – в еритроцитах. Частина вуглекислого газу (від 10 до 20 об’ємних відсотків) може транспортуватися у вигляді сполук з аміногрупою гемоглобіну – карбгемоглобіна.
З усієї кількості СО2 велика його частина (2/3) переноситься плазмою крові.
Однієї з найважливіших реакцій, що забезпечують транспорт СО2, є утворення вугільної кислоти із СО2 і Н2О:
Н2О + СО2 = Н2 СО3
Така реакція в крові прискорюється приблизно в 20 000 разів. Велика швидкість цієї реакції забезпечується ферментом карбоангідразою. При збільшенні вмісту СО2 в крові (що буває в тканинах) фермент сприяє гідратації СО2 і реакція йде у бік утворення Н2СО3. При зменшенні парціальної напруги СО2 у крові (що має місце в легенях) фермент карбоангідраза сприяє дегідратації Н2СО3 і реакція йде убік утворення СО2 і Н2О. Це забезпечує найбільш швидку віддачу СО2 в альвеолярне повітря.
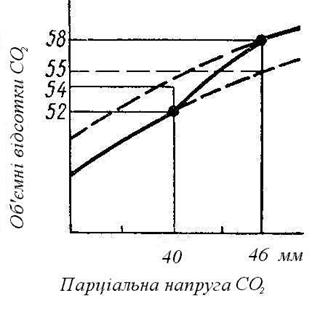 Рис. 91 Криві дисоціації СО2 у крові (по Христіансену, Дугласу і Холдену):
Рис. 91 Криві дисоціації СО2 у крові (по Христіансену, Дугласу і Холдену):
нижня крива – зв’язки СО2 артеріальною кров’ю, гемоглобін якої майже цілком насичений киснем; верхня крива – зв’язки СО2 венозною кров’ю.
Зв’язки СО2 кров’ю, так само як і кисню, залежить від парціального тиску. Можна побудувати криві дисоціації вуглекислоти, відклавши на осі абсцис парціальний тиск СО2, а на осі ординат – кількість пов’язаного вуглекислого газу в об’ємних відсотках.
Крива показує, що зв’язки СО2 кров’ю збільшується в міру зростання його парціального тиску.
При парціальній напрузі CO2, рівній 40 мм рт. ст. (що відповідає його напрузі в артеріальній крові), у крові міститься 52% вуглекислоти. При напрузі CO2, рівній 46 мм рт. cm. (що відповідає напрузі у венозній крові), вміст CO2 зростає до 58%.
На зв’язки CO2 кров’ю впливає присутність оксигемоглобіну в крові. Цю залежність можна простежити при переході артеріальної крові у венозну. Порівняння нижньої кривої і верхньої малюнка 55 показує, що при перетворенні артеріальної крові у венозну солями гемоглобіну віддається кисень і тим самим полегшується її насичення вуглекислим газом. При цьому вміст CO2, в ній збільшується на 6%: з 52% до 58%.
У судинах легень утворення оксигемоглобіну сприяє віддачі CO2, вміст якого при перетворенні венозної крові в артеріальну зменшується з 58 до 52 об’ємних відсотків. У присутності кисню з крові віддаляється весь CO2 при його нульовій напрузі в навколишнім середовищі. У присутності азоту, навіть при нульовій напрузі CO2 у навколишнім середовищі, частина його залишається зв’язаним із кров’ю.
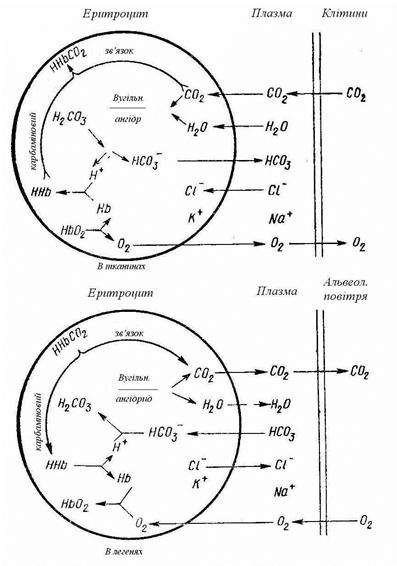 Рис. 92 Транспорт газів. Схема процесів, що відбуваються в еритроциті, при поглинанні чи віддачі кров’ю кисню і вуглекислого газу.
Рис. 92 Транспорт газів. Схема процесів, що відбуваються в еритроциті, при поглинанні чи віддачі кров’ю кисню і вуглекислого газу.
ОБМІН ГАЗІВ У ЛЕГЕНЯХ І ТКАНИНАХ
Обмін газів у легенях. У легенях відбувається обмін газів між альвеолярним повітрям і кров’ю через стінки плоского епітелію альвеол і кровоносних судин. Цей процес залежить від парціального тиску газів в альвеолярному повітрі і їх напруги в крові.
Оскільки парціальний тиск О2 в альвеолярному повітрі великий, а у венозній крові його напруга значна менше, то О2 дифундує з альвеолярного повітря в кров, а вуглекислий газ, унаслідок його більшої напруги у венозній крові, переходить з її в альвеолярне повітря. Дифузія газів здійснюється до настання рівності парціальних тисків. При цьому венозна кров перетворюється в артеріальну – вона одержує 7 об’ємних відсотків кисню і віддає 6 об’ємних відсотків вуглекислого газу.
Кожен газ, перш ніж перейти в зв’язаний стан, знаходиться в стані фізичного розчинення. Кисень, пройшовши цю фазу, надходить в еритроцит, де з’єднується з гемоглобіном і перетворюється в оксигемоглобін:
ННЬ + О2 ® ННЬО2
Оскільки оксигемоглобін є більш сильною кислотою, чим вугільна, то він в еритроцитах реагує з бікарбонатом калію, унаслідок чого утвориться калійна сіль оксигемоглобіну –KHbO2) і вугільна кислота:
КHCO3 + НHbO2 ® KhbO2 + Н2СO3
Утворена вугільна кислота під впливом карбоангідрази піддається дегідратації: H2CO3 ® Н2О + СО2 і вуглекислий газ, що утвориться, виділяється в альвеолярне повітря.
В міру зменшення вуглекислоти в еритроциті на зміну їй із плазми крові надходять іони HCO3-, що утворяться внаслідок дисоціації бікарбонату натрію: NaHCO3 ® Na+ + НСО3-. Замість іонів НСО3- з еритроцитів у плазму надходять іони Cl.
Обмін газів у тканинах. Артеріальна кров, що приходить до тканин, містить 19 об’ємних відсотків кисню, парціальна напруга якого дорівнює 100 мм рт. ст., і 52 об’ємних відсотка CO2 з напругою 40 мм рт. ст.
Оскільки в тканинах у процесі обміну речовин кисень безупинно використовується, то його напруга в тканинній рідині утримується біля нуля. Тому О2 в силу різниці напруг дифундує з артеріальної крові в тканини.
У результаті обмінних процесів, що відбуваються в тканинах, утвориться CO2 і його напруга в тканинній рідині дорівнює 60 мм рт. ст., а в артеріальній крові значно менше. Тому CO2 дифундує з тканин у кров у бік меншої напруги. Вуглекислий газ, надходячи з тканинної рідини в плазму крові, приєднує воду і перетворюється на слабку, легко дисоціюючу вугільну кислоту: Н2О + CO2 = Н2СОз. Н2СО3 диссоціює на іони Н+ і НСО3-: Н2СОз ® Н+ + НСОз-, і її кількість зменшується, унаслідок чого підсилюється утворення Н2СОз із CO2 і Н2О, що поліпшує зв’язки вуглекислого газу. У цілому при цьому зв’язується невелика кількість CO2, тому що константа дисоціації Н2СО3 невелика. Зв’язки CO2, головним чином забезпечують білки плазми крові. Білки, володіючи амфотерними властивостями, у плазмі крові поводяться як слабкі кислоти і утворюють солі з лужними металами. Вступаючи в реакцію з вугільною кислотою, вони здійснюють буферний ефект: аніон білка зв’язує водневі іони, утворити недисоційовану молекулу (зменшує кислотність середовища), а в дисоційованому стані залишається аніон вугільної кислоти і катіон металу.
Н+ + НСО3 + B+ + Р- ® HP + НСО3- + В+
(В+ – катіон металу, Р- – білковий аніон). Катіон металу й аніон НСО3- утворять диссоциирующий бікарбонат: НСО3- +В+ = ВНСО3. Зв’язки водневих іонів і утворення бікарбонатів за допомогою білків забезпечує постійний перенос вуглекислого газу від тканин до легень.
Ведучу роль у переносі вуглекислого газу грає білок гемоглобін. Оболонка еритроцита проникна для вуглекислого газу, що, потрапляючи в еритроцит, під впливом карбоангідрази піддається гідратації і перетворюється в Н2СО3. У капілярах тканин калієва сіль оксигемоглобіну (KНbO2),взаємодіючи з вугільною кислотою, утворить бікарбонат калію (КНСО3), відновлений гемоглобін (ННb) і кисень, що віддається тканинам.
Одночасно вугільна кислота дисоціює: Н2СО3 ® Н+ +НСО3-.Концентрація іонів НСО3- в еритроцитах стає більшою, ніж у плазмі, і вони з еритроцита переходять у плазму. У плазмі аніон НСО3- зв’язується з катіоном натрію Na+ і утвориться бікарбонат натрію (NaНСОз). З плазми крові замість аніонів НСО3- в еритроцити переходять аніони Cl-. Так відбувається зв’язки СО2, що надходить у кров із тканин і перенос його до легень. СО2 переноситься в основному у вигляді бікарбонату натрію в плазмі і частково у вигляді бікарбонату калію в еритроцитах.
РЕГУЛЯЦІЯ ДИХАННЯ
У процесі нервової і гуморальної регуляції дихання забезпечується узгоджена діяльність дихальних м’язів і ритмічна зміна актів вдиху і видиху.
Нервова регуляція дихання здійснюється різними відділами головного мозку, імпульси від яких передаються в спинний мозок, де розташовані нейрони, що регулюють дихальну мускулатуру. У III-V шийних сегментах спинного мозку розташовуються клітини, що регулюють діяльність діафрагми, а в передніх рогах грудних (торакальних) сегментів – клітини, що регулюють міжреберну мускулатуру.
Дихальний центр. Під дихальним центром розуміють сукупність нейронів, розташованих у різних відділах центральної нервової системи, що забезпечують координовану діяльність м’язів і пристосування дихання до мінливих умов зовнішнього і внутрішнього середовища організму.
Н. Д. Миславский (1881) показав, що дихальний центр розташовується в ядрах ретикулярної формації на дні IV шлуночка і захоплює кілька міліметрів в обидва боки. Дихальний центр являє собою парне утворення, що складається з центра вдиху (інспіраторного) і центра виходу (експіраторного). Кожен центр регулює дихання однойменної сторони: при руйнуванні дихального центра з однієї сторони настає припинення дихальних рухів з цієї ж сторони. Миславский висловив думку про те, що дихальний центр є збирачем рефлекторних впливів від усіх чутливих нервів, які впливають на ритм дихання і координацію дихальних рухів.
Значно пізніше кількома дослідниками було показано, що в регуляції акта дихання беруть участь і інші відділи центральної нервової системи.
Нейрони верхнього відділу моста, що регулюють акт дихання, були названі пневмотаксичним центром.
В даний час показано наявність експіраторних та інспіраторних нейронів у довгастому мозку і ретикулярній формації стовбура. Вони розташовані між задньою границею ядер лицьових нервів і місцем відходження корінців від перших сегментів спинного мозку (протягом 10-12 мм). Нейрони експіраторного та інспіраторного центрів розташовуються дифузно. У центрі вдиху зустрічаються нейрони, що регулюють видих, а в центрі видиху – регулюючі вдих.
У структурах моста розрізняють теж два дихальних центри. Один з них – пневмотаксический – сприяє зміні вдиху на видих (гальмує вдих). Другий центр здійснює тонічний вплив на дихальний центр довгастого мозку.
Рис. 93 Локалізація дихальних центрів у нижній частині стовбура мозку (вид позаду):
ПН – пневмотаксичний центр; ІНСП – інс-піраторний;
ЕКСП – експіраторний. Центри є двосторонніми, але для спрощення схеми на кожній із сторін зображений тільки один. Перереэка по лінії 1 не відбивається на диханні, по лінії 2 відокремлюється пневмотаксичний центр, нижче лінії 3 настає зупинка дихання.
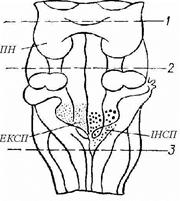
Експіраторний і інспіратор ный центри знаходяться в реципрокных отношениях. Під впливом спонтанної активності нейронів інспіраторного центра виникає акт вдиху, під час якого при розтяганні легень збуджуються механорецептори. Імпульси від механорецепторів по аферентних волокнах блукаючого нерва надходять у дихальний центр і викликають порушення експіраторного і гальмування інспіраторного центра. Це забезпечує зміну вдиху на видих.
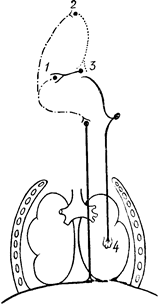 Рис. 94. Схема нервових зв’язків дихального центра; 1 – інспіраторний центр;
Рис. 94. Схема нервових зв’язків дихального центра; 1 – інспіраторний центр;
2 – пневмотаксичний центр;
3 – експіраторний центр;
4 – механорецептори легені.
У зміні вдиху на видих істотне значення має пневмотаксический центр, що свої впливи здійснює через нейрони експіраторного центра. У момент порушення інспіраторного центра довгастого мозку одночасно виникає порушення в інспіраторному відділі пневмотаксичного центра. Від останнього по відростках його нейронів імпульси приходять до експіраторного центра довгастого мозку, викликаючи його порушення і по індукції гальмування інспіраторного центра, що приводить до зміни вдиху на видих.
Таким чином, регуляція дихання здійснюється завдяки узгодженій діяльності усіх відділів центральної нервової системи, об’єднаних поняттям дихального центра. На ступінь активності і взаємодії відділів дихального центра впливають різні гуморальні і рефлекторні фактори.
 Автоматія дихального центра. Здатність дихального центра до автоматії уперше виявлена І. М. Сєченовим (1882 р.) у дослідах на жабах в умовах повної деаферентації тварин. У цих експериментах, незважаючи на те що аферентні імпульси не надходили в центральну нервову систему, реєструвалися коливання потенціалів у дихальному центрі довгастого мозку.
Автоматія дихального центра. Здатність дихального центра до автоматії уперше виявлена І. М. Сєченовим (1882 р.) у дослідах на жабах в умовах повної деаферентації тварин. У цих експериментах, незважаючи на те що аферентні імпульси не надходили в центральну нервову систему, реєструвалися коливання потенціалів у дихальному центрі довгастого мозку.
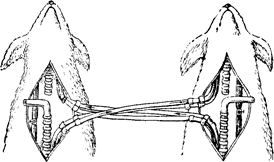
Рис. 95. Схема досліду Фредеріка
з перехресним кровообігом.
Про автоматію дихального центру говорить дослід Гейманса з ізольованою головою собаки. Її мозок був перерізаний на рівні середини моста і позбавлений різних афферентных впливів (були перерізані язикоглотковий, язичний і трійчастий нерви). У цих умовах до дихального центра не надходили імпульси не тільки від легень і дихальних м’язів (унаслідок попереднього відділення голови), але і від верхніх дихальних шляхів (унаслідок перерізання вище названих нервів). Але у тварини зберігалися ритмічні рухи гортані. Цей факт можна пояснити тільки наявністю ритмічної активності нейронів дихального центра.
Автоматия дихального центра підтримується і змінюється під впливом імпульсів від дихальних м’язів, судинних рефлексогенних зон, різних інтеро- і екстерорецепторів, а також під впливом багатьох гуморальних факторів (рН крові, вмісту CO2 і кисню в крові та ін.).
При перфузії IV шлуночка зрушення рН перфузійного розчину в кислу сторону приводить до збільшення легеневої вентиляції, а зрушення рН у лужну сторону – до її зменшення.
Наступними дослідженнями ряду авторів було показано, що в регуляції дихання важливо не тільки зміна рН крові, але і вміст вуглекислоти, як специфічно діючого фактора.
Вплив СО2 на стан дихального центра. Вплив СО2 на активність дихального центра особливо яскраво демонструється в досліді Фредеріка з перехресним кровообігом. У двох собак перерізають сонні артерії і яремні вени і з’єднують перехресно: периферичний кінець сонної артерії з’єднують з центральним кінцем цієї ж судини другого собаки. Так само перехресно з’єднуються і яремні вени: центральний кінець яремної вени першого собаки з’єднується з периферичним кінцем яремної вени другого собаки. У результаті цього кров від тулуба першого собаки надходить до голови другого собаки, а кров від тулуба другого собаки надходить до голови першого собаки. Всі інші судини перев’язують.
Після такої операції в першого собаки робили затиснення трахеї (удушення). Це приводило до того, що через якийсь час спостерігалося збільшення глибини і частоти дихання в другого собаки (диспное), тоді як у першого собаки наставала зупинка дихання (апное). Пояснюється це тим, що в першого собаки в результаті затиснення трахеї не здійснювався обмін газів і в крові збільшувався вміст СО2 (наставала гіперкапнія) і зменшувався вміст кисню. Ця кров надходила до голови другого собаки і впливала на клітини дихального центру, наслідком чого було диспное. Але в процесі посиленої вентиляції легень у крові другого собаки зменшувався вміст СО2 (гіпокапнія) і збільшувався вміст О2. Кров із зменшеним вмістом СО2 надходила до клітин дихального центра першого собаки, і подразнення останнього зменшувалося, що приводило до апное.
Таким чином, збільшення вмісту CO2 у крові приводить до збільшення глибини і частоти дихання, а зменшення змісту СО2 і збільшення О2 – до його зменшення аж до зупинки дихання. У тих спостереженнях, коли першому собаці давали дихати різними газовими сумішами, найбільша зміна дихання спостерігалося при збільшенні вмісту СО2 у крові.
Вплив на дихання вмісту вуглекислого газу у вдихуваному повітрі було показано Холденом у дослідах на людині. Людину поміщали в невелику герметично зачинену камеру, поступово в нього виникало диспное внаслідок збільшення вмісту СО2, у вдихуваному повітрі. У цих же дослідах було показано, що зменшення вмісту кисню у вдихуваному повітрі не впливає на дихання: коли з камери поглиначем витягали вуглекислий газ, диспное не наставало. Зі збільшенням вмісту СО2 у вдихуваному повітрі зростає легенева вентиляція.
Залежність величини легочнол вентиляції від вмісту СО2, у вдихуваному і альвеолярному повітрі (у %)
| Вміст СО2 у повітрі | Легенева вентиляція | |
| у вдихуваному | у альвеолярному | |
| 0,03 | 5,71 | |
| 3,98 | 6,03 | |
| 5,28 | 6,53 |
В даний час установлено, що при збільшенні змісту О2 в крові на 0,2% легенева вентиляція зростає на 100%.
Про вплив вмісту CO2 у крові на дихання говорять і найпростіші досліди з гіпо- і гіпервентиляцією легень. Якщо людина довільно затримає дихання, то через якийсь час затримка змінюється глибоким і частим диханням. Це відбувається внаслідок того, що під час затримки дихання в крові накопичується CO2. Кров зі збільшеним вмістом вуглекислоти омиває клітини дихального центра і стимулює його діяльність.
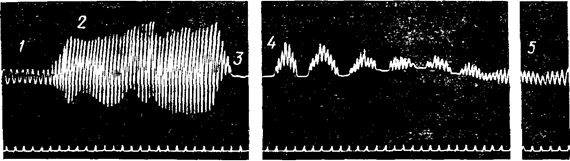 |
Рис. 96 Ефект гіпервентиляції легень:
1 – нормальне дихання; 2 – довільне посилення дихання
(максимальна глибина і частота) протягом двох хвилин;
3 – апное протягом двох хвилин; 4 – нерівномірне дихання (чейн-стоксове);
5 – нормальне дихання, записане через декілька хвилин; унизу – оцінка часу.
Якщо людина буде робити глибокі вдихи і видихи (гіпервентиляцію легень), то через якийсь час настає короткочасна затримка дихання.
Після затримки дихання воно стає нерівним, а його амплітуда і частота трохи зменшуються в порівнянні з вихідним рівнем, що має місце до гіпервентиляції. Ці зміни дихання зв’язані з тим, що під час гіпервентиляції легень із крові видаляється велика кількість CO2, що викликає падіння активності дихального центра.
При вдиханні газової суміші з підвищеним вмістом CO2 у людини настає різке збільшення глибини дихальних рухів.
Механізм впливу СО2 на дихальний центр. Останнім часом показана присутність хеморецепторів у ретикулярній формації мозкового стовбура. Вони являють собою тільця величиною близько 2 мм, що розташовуються з двох сторін від дихального центра на вентролатеральній поверхні довгастого мозку, поблизу від місця виходу під’язичного нерва. Установлено, що мембрана хеморецепторних клітин має підвищену проникність до CO2, після проникнення якого у клітку відбуваються утворення HgCOs і наступна дисоціація її на іони Н4" і НСОГ. Водневі іони, що утворяться, є подразником хеморецепторов.
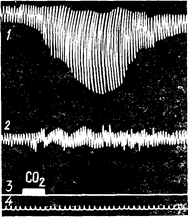
Рис. 97 Ефект вдихання повітря,
що містить 6% вуглекислого газу;
1 – запис дихальних рухів; 2 ¾ ков’яного тиску;
3 – тривалості вдихання газу; 4 – оцінка часу.
Саме швидкою дифузією CO2 через мембрану хеморецепторних клітин пояснюється вибірковість дії вугільної кислоти на дихальний центр.
При збільшенні вмісту кисню в крові частота і глибина дихання зменшуються, а при зменшенні його вмісту – збільшуються. Це вплив значною мірою пов’язаний з рефлекторним впливом кисню на клітини дихального центра через хеморецептори судинних рефлексогенних зон.
РЕФЛЕКТОРНА РЕГУЛЯЦІЯ ДИХАННЯ
Рефлекторні впливи на дихання із судинних рефлексогенних зон. Дихання особливо швидко реагує на різні подразнення. Воно швидко змінюється під впливом імпульсів, що приходять з екстеро- і інтерорецепторів до клітин дихального центра.
Подразником рецепторів можуть бути хімічні, механічні, температурні й інші впливи. Найбільше яскраво вираженим механізмом саморегуляції є зміна дихання під впливам хімічного і механічного подразнення судинних рефлексогенних зон, механічного подразнення рецепторів легень і дихальних м’язів.
Різними дослідниками було показано, що зміна хімічного складу крові впливає не тільки прямим шляхом на стан дихального центра, але і рефлекторно, насамперед через судинні рефлексогенні зони.
Синокаротидна судинна рефлексогенна зона містить рецептори, чутливі до вмісту CO2, O2 і водневих іонів у крові. Це чітко показано в дослідах Гейманса з ізольованим каротидним синусом, що відокремлювали від сонної артерії і постачали кров’ю від іншої тварини. З центральною нервовою системою каротидный синус був з’єднаний тільки нервовим шляхом – зберігався нерв Геринга. При підвищенні вмісту СО2 у крові, що омиває каротидне тільце, виникає порушення хеморецепторов цієї зони, унаслідок чого збільшується кількість імпульсів, що йдуть до дихального центра, і настає рефлекторне збільшення глибини дихання.
Збільшення глибини дихання настає і під впливом СО2 на хеморецептори аортальної рефлексогенної зони.
Такі ж зміни дихання настають при подразненні хеморецепторов названих рефлексогенних зон кров’ю з підвищеною концентрацією водневих іонів.
У тих же випадках, коли в крові збільшується вмістO2,подразнення хеморецепторов рефлексогенних зон зменшується, унаслідок чого слабшає потік імпульсів до дихального центра і настає рефлекторне зменшення частоти дихання.
Інші зміни мають місце при збіднінні крові О2 (гіпоксемії). У дослідах Гейманса показано, що зменшення вмісту О2 у крові є подразником хеморецепторов каротидного тельця. При реєстрації потенціалів із синокаротидного нерва було виявлено їх частішання у випадку перфузії каротидного синуса кров’ю зі зниженим вмістом O2. Одночасно наставало збільшення частоти дихання. Ці зміни не настають після руйнування каротидного тільця чи його денервації.
Рефлекторним збудником дихального центра і фактором, що впливає на дихання, є зміна кров’яного тиску в судинних рефлексогенних зон. При підвищенні артеріального тиску подразнюються механорецептори судинних рефлексогенних зон, унаслідок чого настає рефлекторне пригнічення дихання. Зменшення величини артеріального тиску приводить до збільшення глибини і частоти дихання.
Рефлекторні впливи на дихання з механорецепторов легень і дихальних м’язів. Істотним фактором, що викликає зміну вдиху і видиху, є впливи з механорецепторов легень, що вперше було виявлено Герингом і Брейером (1868 р.). Вони показали, що кожен вдих стимулює видих. Під час вдиху, при розтяганні легень, подразнюються механорецепторы, розташовані в альвеолах і дихальних м’язах. Виниклі в них імпульси по аферентним волокнам блукаючого і міжреберних нервів приходять до дихального центра і викликають гальмування інспіраторних нейронів і порушення експіраторних, викликаючи зміну вдиху на видих. Це один з механізмів саморегуляції дихання.
Отримані електрофізіологічні докази рефлекторних впливів на дихальний центр із механорецепторів легень. Записуючи потенціали з периферичних кінців перерізаних блукаючих нервів, можна бачити, що при звичайному природному диханні в момент вдиху виникають часті потенціали. При спокійному пасивному видиху вони відсутні, тому що механорецептори легень не подразнюються. Тільки при дуже глибокому видиху (при сильному спадінні легень) виникає подразнення механорецепторів, що стає однією з причин зміни видиху на вдих.
Дослідженнями останніх років показано, що рефлекторні впливи з механорецепторів легень можуть передаватися і не по блукаючих нервах. Вони можуть безпосередньо поширюватися по еферентним спинальним нейронам по аферентним нервам, що йде від легень у складі задніх корінців спинного мозку. Цей механізм важливий при глибокому диханні під час різних фізичних (особливо спортивних) навантажень.
Подібно рефлексу Геринг – Брейера здійснюються рефлекторні впливи на дихальний центр від рецепторів діафрагми. Під час вдиху в діафрагмі при скороченні її м’язових волокон подразнюються закінчення нервових волокон, імпульси що виникли в них надходять у дихальний центр і викликають припинення вдиху і виникнення видиху. Цей механізм має особливо велике значення при посиленому диханні.
Рефлекторні впливи на дихання з різних рецепторів організму. Розглянуті вище рефлекторні впливи на дихання відносяться до постійно діючих. Але існують різні короткочасні впливи майже з усіх рецепторів нашого організму, що впливають на дихання.
Так, при дії механічних і температурних подразників на екстерорецептори шкіри настає затримка дихання. При дії холодної чи гарячої води на велику поверхню шкіри виникає зупинка дихання на вдиху. Болюче подразнення шкіри викликає різкий вдих (скрикування при болі являє собою вдих з одночасним закриттям голосової щілини).
На характер дихання впливають імпульси з рецепторів травного апарата. Такі фізіологічні реакції, як акт ковтання, блювоти й акт дефекації, супроводжуються зміною дихання.
Деякі зміни акта дихання, що виникають при подразненні слизових оболонок дихальних шляхів, одержали назву захисних дихальних рефлексів. До них відносяться такі рефлекторні акти, як кашель, чихання, затримка дихання, що настає при дії різких запахових подразників і ін.
РОЛЬ КОРИ ВЕЛИКИХ ПІВКУЛЬ ГОЛОВНОГО МОЗКУ В РЕГУЛЯЦІЇ ДИХАННЯ
Дихання – це одна з вегетативних функцій, що має довільну регуляцію. Кожна людина може довільно змінити ритм і глибину дихання, затримати його на визначений час (10-20 сек). Можливість довільної зміни дихання говорить про регулюючі впливи кори великих півкуль на дану функцію.
Перші експериментальні докази кіркової регуляції дихання були представлені В. Я. Данилевським (1874 р.). Він показав, що при подразненні моторної ділянки кори виникають зміни дихання, характер яких залежить від місця й інтенсивності подразнення. У подальшому багато вчених у дослідах з подразненням і екстирпацією різних відділів кори великих півкуль підтвердили факт кіркової регуляції дихання.
У зв’язку з цим тривалий час обговорювалося питання про існування точної локалізації дихального центра в корі великих півкуль. В даний час вважають, що на дихання впливають різні відділи кори великих півкуль, але подразнення деяки
Читайте також:
- I. Особливості аферентних і еферентних шляхів вегетативного і соматичного відділів нервової системи
- II. Мотивація навчальної діяльності. Визначення теми і мети уроку
- IV. НС у природному середовищі та інших сферах життєдіяльності людини
- IІI. Формулювання мети і завдань уроку. Мотивація учбової діяльності
- V. Питання туристично-спортивної діяльності
- VI.3.3. Особливості концепції Йоганна Гайнріха Песталоцці
- VI.3.4. Особливості концепції Йоганна Фрідриха Гербарта
- А. Особливості диференціації навчального процесу в школах США
- Абсолютні та відності показники результатів діяльності підприємства.
- Автоматизація банківської діяльності в Україні
- Автоматизація метрологічної діяльності
- Агітація за і проти та деякі особливості її техніки.
| <== попередня сторінка | | | наступна сторінка ==> |
| Зміна частоти серцевих скорочень у дітей з віком (за А. Ф. Туру) | | | Зміна величини дихального об’єму легень з віком |
|
Не знайшли потрібну інформацію? Скористайтесь пошуком google: |
© studopedia.com.ua При використанні або копіюванні матеріалів пряме посилання на сайт обов'язкове. |