РЕЗОЛЮЦІЯ: Громадського обговорення навчальної програми статевого виховання
ЧОМУ ФОНД ОЛЕНИ ПІНЧУК І МОЗ УКРАЇНИ ПРОПАГУЮТЬ "СЕКСУАЛЬНІ УРОКИ"
ЕКЗИСТЕНЦІЙНО-ПСИХОЛОГІЧНІ ОСНОВИ ПОРУШЕННЯ СТАТЕВОЇ ІДЕНТИЧНОСТІ ПІДЛІТКІВ
Батьківський, громадянський рух в Україні закликає МОН зупинити тотальну сексуалізацію дітей і підлітків
Відкрите звернення Міністру освіти й науки України - Гриневич Лілії Михайлівні
Представництво українського жіноцтва в ООН: низький рівень культури спілкування в соціальних мережах
Гендерна антидискримінаційна експертиза може зробити нас моральними рабами
ЛІВИЙ МАРКСИЗМ У НОВИХ ПІДРУЧНИКАХ ДЛЯ ШКОЛЯРІВ
ВІДКРИТА ЗАЯВА на підтримку позиції Ганни Турчинової та права кожної людини на свободу думки, світогляду та вираження поглядів
- Гідрологія і Гідрометрія
- Господарське право
- Економіка будівництва
- Економіка природокористування
- Економічна теорія
- Земельне право
- Історія України
- Кримінально виконавче право
- Медична радіологія
- Методи аналізу
- Міжнародне приватне право
- Міжнародний маркетинг
- Основи екології
- Предмет Політологія
- Соціальне страхування
- Технічні засоби організації дорожнього руху
- Товарознавство продовольчих товарів
Тлумачний словник
Авто
Автоматизація
Архітектура
Астрономія
Аудит
Біологія
Будівництво
Бухгалтерія
Винахідництво
Виробництво
Військова справа
Генетика
Географія
Геологія
Господарство
Держава
Дім
Екологія
Економетрика
Економіка
Електроніка
Журналістика та ЗМІ
Зв'язок
Іноземні мови
Інформатика
Історія
Комп'ютери
Креслення
Кулінарія
Культура
Лексикологія
Література
Логіка
Маркетинг
Математика
Машинобудування
Медицина
Менеджмент
Метали і Зварювання
Механіка
Мистецтво
Музика
Населення
Освіта
Охорона безпеки життя
Охорона Праці
Педагогіка
Політика
Право
Програмування
Промисловість
Психологія
Радіо
Регилия
Соціологія
Спорт
Стандартизація
Технології
Торгівля
Туризм
Фізика
Фізіологія
Філософія
Фінанси
Хімія
Юриспунденкция
Відмінності гуморальної регуляції від нервової
| Гуморальна регуляція | Нервова регуляція |
| носій інформації | |
| хімічна речовина | потенціал дії |
| характер передавання інформації | |
| кровоносні й лімфатичні судини та міжклітинні простори | нервові волокна; |
| спосіб передавання інформації | |
| потік крові, лімфи та дифузія | поширення потенціалу дії нервовими волокнами |
| Швидкість поширення сигналу | |
| невеликою швидкістю | швидко |
Гуморальний спосіб передавання інформації має такі особливості:
¾ гуморальний сигнал можна менш точно дозувати за силою і тривалістю, ніж збудження, що пояснюється повільним зростанням концентрації гуморального агента, причому не до суворо визначеної певної величини, і так само його поступовим зменшенням;
¾ гуморальна регуляція використовується в організмі для забезпечення реакцій, які не потребують терміновості й точності.
¾ Гуморальна регуляція, як і нервова, характеризується певною направленістю за принципом "усім, усім, хто відізветься". При цьому направленість гуморальної регуляції обумовлена тим (здійснюється "за точною адресою"), що лише певні клітини, "органи-мішені", чутливі до якого-небудь гуморального фактора, які мають відповідні рецептори.
Це можна зрозуміти на прикладі, коли радіопередачі транслюються за допомогою радіохвиль в ефір для всіх. Водночас приймають їх лише радіоприймачі, налаштовані на хвилю відповідної довжини.
У нервовій системі збудження передається нервовими волокнами "за точною адресою" і без спотворення ("передача телеграфними проводами").
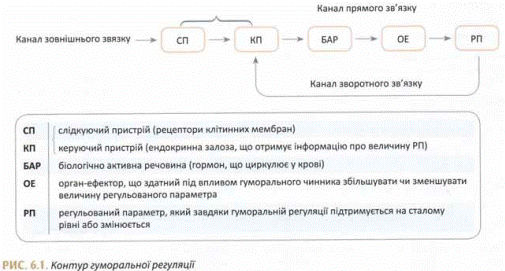
Гуморальна регуляція, як і нервова, завжди здійснюється замкнутими контурами. Однак у таких контурах функції керуючого пристрою виконує сама секреторна клітина.
Обов'язковими ланками контуру гуморальної регуляції є:
а) регульований параметр;
б) виконавчі механізми або органи;
в) канали прямого і зворотного зв'язку.
Факторами гуморальної регуляції (первинними месенджерами, або посередниками) є:
а) гормони;
б) гормоноїди (тканинні або місцеві гормони);
в) деякі метаболіти й іони (виділяють справжні метаболіти й фактори пошкодження).
2.Ендокринні залози, їх види, гормони та значення.
Умовно серед елементів ендокринної системи організму розрізняють чотири групи складників: центральні, периферійні, органи із змішаними функціями та дисоційовані ендокринні клітини.
1. Центральні органи ендокринної системи — гіпоталамус, гіпофіз і епіфіз. Ці органи тісно пов'язані з органами центральної нервової системи і координують діяльність усіх інших ланок ендокринної системи.
2. Периферійні ендокринні органи— щитовидна, прищитовидні і надниркові залози. Це суто ендокринні залози, які здійснюють багатоплановий вплив на організм, посилюючи або послаблюючи обмінні процеси.
3. Органи, які поєднують виконання ендокринної функції з рядом інших - підшлункова залоза, статеві залози (яєчко, яєчник), нирки, плацента тощо.
4. Дисоційована ендокринна система — ендокриноцити, які лежать в різних органах поодинці, можуть мати епітеліальне походження або нейральне. Останні здатні захоплювати з крові та декоарбоксилювати попередники амінів, тому належать до так званої APUD-системи (від англ. amine precursors uptake and decarboxylation). Ці клітини виявляються при імпрегнації сріблом, тому ще називаються аргентафінними.
Залозами внутрішньої секреції, або ендокринними залозами, є спеціалізовані органи, які виділяють утворені в них продукти секреції безпосередньо в кров або тканинну рідину. В даний час до ендокринних залоз відносять гіпофіз, щитовидну залозу, паращитовидні залози, кіркова і мозкова речовина надниркових залоз, острівцевий апарат підшлункової залози, статеві залози, тимус, і епіфіз. Ендокринної активністю володіє також плацента. Крім того, ендокринні клітини можуть присутні в деяких інших органах і тканинах, зокрема в травному тракті, нирках, серцевому м'язі, вегетативних гангліях. Ці клітини утворюють так звану дифузну ендокринну систему. Загальною функцією для всіх залоз внутрішньої секреції є вироблення гормонів.
| Залоза. | Гормони. | Дія гормонів на організм. |
| Аденогіпофіз. | Соматотропний (СТГ) | Посилює лінійний ріст і масу тіла організму (посилює протеосинтез і ліполіз). |
| Тиреотропний (ТТГ) | Викликає виділення гормонів щитовидної залози. | |
| Адренокортикотропний (АКТГ) | Викликає виділення гормонів кіркового шару наднирників, перш за все глюкокортикоїдів. | |
| Гонадотропні (фолікулостимулюючий, лютеїнізуючий). | Викликають виділення статевих гормонів. | |
| Пролактин. | Стимулює утворення та виділення молока молочними залозами. | |
| Меланоцитостимулюючий. | Стимулює утворення меланіну (пігмента шкіри). | |
| Нейрогіпофіз. | Вазопресин. | Підвищує реабсорбцію води в дистальних канальцях нефрона. |
| Окситоцин. | Стимулює скорочення м’язів матки при родах. | |
| Щитовидна залоза. | Трийодтиронін (Т3). Тетрайодтиронін (Т4 або тирозин). | Посилюють окисний метаболізм; забезпечують диференціювання структур головного мозку у ново-народжених. |
| Тиреокальцитонін. | Зменшує вміст іонів кальцію в крові через збільшення надходження його в кісткову тканину. | |
| Паращитовидні залози. | Паратгормон (ПТГ) | Підвищують вміст іонів кальцію в крові через посилення його реабсорбції в нирках, кишківнику та резорбції з кісткової тканини. |
| Наднирники (мозковий шар). | Катехоламіни (адреналін; норадреналін). | Посилюють ЧСС, ССС, розширюють бронхи, гальмують секрецію і моторику ШКТ; викликають гіпер-глікемію та підвищення вмісту жирних кислот в крові. |
| Наднирники (кірковий шар). | Глюкокортикоїди (кортизол). | Викликають гіперглікемію через посилення глюконеогенезу в печінці. |
| Мінералокортикоїди (альдостерон). | Посилюють реабсорбцію іонів натрію і секрецію іонів калію в канальцях нирки. | |
| Статеві гормони. | Стимулюють розвиток вторинних статевих ознак, забезпечуютьвиконання статевих функцій. | |
| Підшлункова залоза. | Інсулін. | Знижує рівень глюкози в крові (через посилення її проникнення в клітини). |
| Глюкагон. | Викликає гіперглікемію шляхом посилення розпаду глікогену в печінці. |
3.Основні види та механізми дії гормонів.
Генералізовані спеціалізовані ефекти гормональної регуляції здійснюються за допомогою особливих хімічних регуляторів внутрішнього середовища - гормонів.
Гормонаминазивають хімічні речовини, що утворюються і виділяються спеціалізованими ендокринними клітинами у внутрішнє середовище організму для регуляції обміну речовин, вегетативних функцій організму та гуморального забезпечення координації й інтеграції процесів життєдіяльності.
Будучи носіями інформації, гормони беруть участь у регуляції функцій організму, адаптують його до постійно змінюваних умов зовнішнього середовища, забезпечують його ріст, розвиток та розмноження.
Синтез гормонів ендокринними клітинами відбувається безперервно, його інтенсивність залежить не тільки від регуляторних сигналів ланок управління, але й від величини секреції. Гормони можуть депонуватися у тих же тканинах, де утворюються. Депонування гормонів ендокринною тканиною може здійснюватися у спеціалізованих гранулах (мозкова речовина надниркових залоз) або у спеціалізованих структурах залози (колоїд фолікулів щитоподібної залози).
Діють гормони так:
· телекриновий ефект - надходять до рідких середовищ організму і переносяться ними до віддалених клітин-мішеней
· паракриновий ефект – діє місцево на сусідні клітини
· автокриновий ефект – дія на саму клітину
Для регуляції функціонування багатьох органів та процесів цей механізм хоч і діє більш повільно, але виявляється ефективнішим, ніж нервова регуляція. Це зумовлено наступним:
а) біологічно активна сполука може надходити до кожної клітини;
б) спектр зазначених регуляторів значно ширший, ніж нейротрансмітерів;
в) їх дія триваліша.
Серед біологічно активних сполук виділяють:
1. Справжні гормони - секретуються залозами внутрішньої секреції (тироксин, інсулін, паратгормон тощо) або спеціалізованими ендокринними клітинами, що розташовані в різних органах (ліберини, статини, гормони травного тракту тощо).
2. Тканинні гормони - секретуються неспеціалізованими клітинами різних органів (гістамін, серотонін, простагландини, енкефаліни тощо).
3. Метаболіти - речовини, що утворюються у результаті обмінних процесів у тканинах (молочна кислота, аденозин, вуглекислий газ, деякі іони тощо).
Специфічну групу гормонів складають гастроінтестинальні гормони і ангіотензин. Ці гормони не є продуктами ендокринних залоз. Перші синтезуються і виділяються в кров інкреторними клітинами, які входять до складу різних залоз шлунково-кишкового тракту. Ангіотензин утворюється безпосередньо в крові в результаті взаємодії одного з білків плазми крові (а2-глобуліна) з активатором - ферментом реніном, що виділяється в кров інкреторними клітинами юкстагломерулярного апарату нирок.
При цьому в різних органах один і той самий гормон спричиняє різні впливи. Наприклад, ангіотензин у надниркових залозах викликає продукцію альдостерону, в ЦНС - являє собою регулятор спраги, у серцево-судинній системі спричиняє скорочення кардіоміоцитів і гладеньких м'язів судин.
Тканинні гормони (гістогормони) мають місцеву - ізокринну дію.
До гормоноїдів відносять перш за все гуморальні фактори, які утворюються в організмі без участі спеціалізованих інкреторних клітин. Це кініни (калідин і брадикінін), простагландини і серотонін, гістамін, а також відносять медіатори.
Простагландини, гістамін і серотонін виділяються в кров різними клітинами організму, для яких інкреторна функція не є ведучою. Кініни (калідин і брадикінін) утворюються безпосередньо в крові з а2-глобуліну під впливом активатора "калікреїну", що виділяється в кров знову ж таки не інкреторними клітинами, а звичайними зовнішньосекреторними клітинами різних залоз травного каналу.
Медіатори виділяються в синапсах. Якщо в синапсі медіатор не встигає повністю зруйнуватись або поглинутись, то він може потрапляти в тканинну рідину і кров, забезпечуючи регуляторну дію на найближчі клітини.
Гастроінтестинальні гормони утворюються в клітинах, розміщених в шлунково-кишковому тракті, які отримали назву АРUD-системи (амін – прекурсор – аптек – декарбексилейшин, тобто, система захоплення і дезамінування попередників амінів). Вони здатні поглинати й накопичувати попередники біогенних амінів з наступним утворенням біологічно активних речовин і поліпептидних гормонів. Інші клітини АРUD-системи також утворюють нейропередавачі і нейротрансмітери. Нейромодулятори (ацетилхолін, адреналін) також синтезуються в нервових клітинах і звільняються нервовими закінченнями. Сюди відносять також гістамін, серотонін і у-аміномасляну кислоту. Вони діють на дуже короткій відстані від місця синтезу й менший час, ніж пептидні, стероїдні, і білкові та тіреоідні гормони.
Пептиди, аміни і нейромедіатори - гідрофільні сполуки і не можуть легко проникати крізь плазматичну мембрану клітини.
Метаболіти - неспецифічні продукти внутрішньоклітинного енергетичного обміну речовин, які постійно виділяють у кров усі клітини організму, звільняючись від їх надлишку.
Деякі метаболіти мають біологічну активність, тобто можуть забезпечити виникнення певних пристосовних реакцій організму (С02, молочна кислота). Рівень біологічної активності метаболітів, порівняно з активністю гормонів, невисокий. Тому їхня регуляторна дія виявляється лише в і разі їх виділення у великих кількостях.
Саме тому поняття "гуморальна регуляція" ширше, ніж "гормональна регуляція", яка здійснюється лише за допомогою гормонів.
Виділяють п'ять видів дії гормонів:
Розрізняють 5 видів фізіологічної дії гормонів на клітини-мішені:
• метаболічна дія. - викликає зміни обміну речовин в тканинах, які в свою чергу лежать в основі зміни функції клітини, тканини чи органа;
• морфогенетична дія - вплив гормонів на процеси формоутворення, диференціювання та росту структурних елементів організму. Ця дія досягається шляхом впливу на генетичний апарат клітини. Прикладом може служити вплив соматотропіну на ріст тіла та внутрішніх органів і вплив статевих гормонів на розвиток вторинних статевих ознак;
• кінетична дія - здатність гормонів запускати діяльність ефектора, включати реалізацію певної функції. Так, наприклад, окситоцин викликає скорочення мускулатури матки, вазопресин включає реабсорбцію води в збиральних трубочках нефрона; при його відсутності цей процес не відбувається;
• коригуюча дія - зміна діяльності органів чи процесів, які проходять і при відсутності гормона. Прикладом коригуючої дії є вплив адрена-
ліну на частоту серцевих скорочень, активація окисно-відновних процесів тироксином. Різновидністю коригуючої дії є нормалізуючий ефект гормонів на рівень тих чи інших метаболітів (гомеостатична дія). Так, глюкокортикоїди викликають розпад білків при надлишковому їх синтезі в організмі;
• реактогенна дія - здатність гормону змінювати чутливість тканин до дії інших гормонів чи біологічно активних речовин. Так , наприклад, тиреоїдні гормони підсилюють ефекти катехоламінів, гормон фолікулін стимулює дію прогестерона на слизову оболонку матки. Різновидом реактогенної дії є так звана пермісивна (дозволяюча) дія, яка полягає в тому, що ефекти деяких гормонів реалізуються тільки при наявності іншого гормона. Так, глюкокортикоїди мають пермі-сивну дію по відношенню до катехоламінів, а інсулін має пермісивну дію по відношенню до гормону росту.
Крім того, гормони впливають на імуногенез, гемостаз, здійснюють сенсибілізувальний і десенсибілізувальний впливи.
За хімічною природою усі гормони поділяються на три групи:
1) похідні амінокислот - гормони щитоподібної залози, мозкового шару надниркових залоз, епіфізу;
2) пептидні гормони, прості і складні білки - гіпоталамічні нейропепти-ди, гормони гіпофізу, острівкового апарату підшлункової залози, прищиподібних залоз;
3) стероїдні гормони - гормони кіркового шару надниркових залоз, статевих залоз, гормон ниркового походження - кальцитріол.
Руйнування гормонів відбувається ферментативними системами печінки, легень, мозку, нирок. Інколи гормони гідролізуються в самій клітині-мішені, будучи зв'язаними з рецептором або вільними в лізосомі.
На основі функціональних критеріїв розрізняють три групи гормонів:
1)гормони, які впливають безпосередньо на орган-мішень; ці гормони називають ефекторними,
2)гормони, основною функцією яких є регуляція синтезу і виділення ефекторних гормонів, їх називають тропними;
3)гормони, що синтезуються нервовими клітинами в гіпоталамусі, вони регулюють синтез і виділення гормонів аденогіпофізом, їх називають рилізинг-гормонами.
Загальні властивості гормонів:
1. Специфічність дії. Кожен гормон діє на конкретні фізіологічні системи, органи чи тканини, тобто на ті структури, що містять спеціалізовані рецептори до нього.
2. Дистантна дія. Багато гормонів діють через внутрішнє середовище на органи, які розташовані далеко від місця їх утворення. Проте тканинні гормони, що секретуються без участі спеціалізованих ендокринних клітин, найчастіше проявляють місцеву дію на невеликій відстані від місця їх утворення.
3. Відсутність видової специфічності, за винятком гормону росту і ліпотропіну.
4. Дія зумовлена впливом на клітини-мішені плазматичних мембран.
5. Висока біологічна активність - вплив на функції органів і тканин у дуже невеликих концентраціях.
Засобами транспорту гуморальних агентів є кров, лімфа й міжклітинна рідина.
Гормони циркулюють у крові у вільному стані або у вигляді сполук з форменими елементами крові. Більшість гормонів (близько 80%) переносяться у комплексі зі специфічними білками плазми крові, які вибірково транспортують молекули того чи іншого гормону. Після сполучення з білками гормони переходять у неактивну форму, а також стають захищеними від руйнування ферментами крові.
Концентраціябільшості гормонів у крові непостійна. З одного боку, вона визначається інтенсивністю утворення і секреції гормону залозою, а з іншого - швидкістю його поглинання, руйнування та виведення. Час напіввиведення (Т 1/2) гормонів із крові становить понад 4 доби - у тироксину, кілька годин - у стероїдів, хвилини - в інсуліну, вазопресину, катехоламінів і секунди - у нейромедіаторів та пептидів.
Секреція деяких гормонів є періодичною. Наприклад, рівень гормону росту, АКТГ, має виражені добові коливання, які пов'язані з циклами неспання. А для статевих гормонів характерна не лише добова ритміка утворення і секреції, але й чітко виражена періодика, пов'язана із віковими періодами розвитку, менструальним циклом та циклом "вагітність-пологи".
Далеко не завжди в організмі людини гормони утворюються у кількості, що забезпечує їх нормальне функціонування. Функціональна активність залози може бути підвищеною (гіперфункція) або, навпаки, зниженою (гіпофункція). Ця зміна може бути короткочасною, що зумовлено необхідністю змінити активність відповідної функції організму, або мати більш тривалий чи постійний характер і торкатися змін не тільки функції, але й структури ендокринної залози, що характерно для стану хвороби.
Гормональний ефект залежить не тільки від кількості й властивостей молекул гормонів (правило попереднього стану), а й від реактивності ефектора (визначається числом і властивостями мембранних рецепторів до гормонів).
Це можливо внаслідок координації специфічних функцій органів і систем при тісній взаємодії із ЦНС, активуючи каскад реакцій.
Механізм дії гормонів
Дія гормону (ліганда) на клітину реалізується шляхом взаємодії з рецептором.
Рецептори можуть бути розташовані як на мембрані, так і всередині клітини.
Молекулярний циторецептор - це група молекул цитоплазми або мембрани клітини, які мають вибіркову чутливість до певної хімічної речовини (гуморального фактора, медіатора).
Розрізняють три види молекулярних циторецепторів:
- цитоплазматичних,
- мембранних,
- ядерних.
Відповідно розрізняють три механізми дії хімічних речовин на клітини організму:
1. Дія хімічних речовин на клітини через їхні цитоплазматичні рецептори.
У цьому разі біологічно активна речовина проникає крізь мембрану клітини і з'єднується з її цитоплазматичними рецепторами, вибірково чутливими до цього фактора.
2. Дія хімічних речовин на клітини через їхні ядерні рецептори Гормон може прямо проникати в ядро й там зв'язуватися з рецепторами ядра.
3. Дія хімічних речовин на клітини через їхні мембранні рецептори Такий механізм дії на клітину хімічних речовин властивий речовинам з невеликою молекулярною масою і здатним проникати через клітинну мембрану (стероїдні гормони, гормони щитоподібної залози та інші небілкові гормони).
Регулювання функцій синтезу і секреції гормонів ендокринними залозами здійснюється кількома шляхами:
1. Нейрогенне регулювання здійснюється у двох напрямках:
а) прямий вплив нервової системи на синтез і секрецію гормонів (нейрогіпофіз, мозковий шар надниркових залоз);
б) нервова система регулює гормональну активність опосередковано, змінюючи інтенсивність кровопостачання залози
2. Гуморальне регулювання полягає у безпосередньому впливі на клітини залози концентрації субстрату, рівень якого регулює гормон (зворотний зв'язок). Так, рівень кальцію у крові впливає на секреторну активність прищитоподібної і щитоподібної залоз, де утворюються паратгормон і тирокальцитонін, а концентрація глюкози в крові визначає активність виходу інсуліну з підшлункової залози і т.д.
3. Нейрогуморальне регулювання здійснюється за допомогою гіпоталамо-гіпофізарної системи.
Функція щитоподібної та статевих залоз, кори надниркових залоз регулюється гормонами передньої частки гіпофіза (аденогіпофізом) - тропними гормонами: адренокортикотропним, тиреотропним, фолікулостимулювальним і лютеїнізивним. Дещо умовно до тропних гормонів відносять і соматотропний гормон гіпофіза, що здійснює свій вплив на ріст ще й опосередковано через гормон соматомедин, який утворюється у печінці.
В аденогіпофізі також утворюється інтермедин (меланоцитостимулювальний гормон, МСГ) і пролактин, що здійснюють прямий вплив на периферійні органи.
У свою чергу, вивільнення усіх зазначених гормонів аденогіпофіза залежить від гормональної активності нейронів медіальної ділянки гіпоталамуса. Тут утворюються гормони, що здійснюють стимулювальний чи гальмівний вплив на секрецію гормонів аденогіпофіза: рилізинг-фактори (ліберини) та інгібітори -статики.
Ліберини і статини, впливаючи на продукцію тропних гормонів аденогіпофіза, регулюють активність деяких залоз внутрішньої секреції.
Останнім часом використовують спільну назву - гіпофізіотропні гормони. На сьогодні встановлено сім гіпоталамічних рилізинг - та гальмівних гормонів:
- кортикотропін-рилізинг гормон (КРГ),
- тиротропін-рилізинг-гормон (ТРГ),
- гормон росту - рилізинг гормон (ГРРГ),
- гормон росту інгібувальний гормон (ГРІ, або соматостатин),
- лютеїнізивний гормон -рилізинг гормон (ЛГРГ) або гонадотропін-рилізинг гормон (ГнРГ) ,
- пролактинінгібувальний гормон (ПІГ).
- пролактин-рилізинг-гормон (ПРГ).
Ділянка, в якій відбувається виділення гіпоталамічних рилізинг-гормонів та гальмівних гормонів, є серединним підвищенням гіпоталамуса. Вона містить небагато тіл нервових клітин, проте має чимало нервових закінчень, розміщених безпосередньо біля капілярних петель, з яких починаються портальні судини, що з'єднуються з гіпофізарною портальною системою судин.
Функція значної кількості клітин, що синтезують гормони, регулюється за допомогою комбінації кількох названих механізмів. Крім того, багато гормонів, а також і гіпоталамо-гіпофізарні фактори впливають на утворення один одного. Тому, оцінюючи той чи інший механізм регуляції, необхідно мати на увазі, що мова йде лише про переважання того чи іншого з них.
У цілому вони беруть участь у забезпеченні гомеостазу; адаптації організму до різних умов існування та швидкої їх зміни; регуляції процесів росту; фізичного, статевого й інтелектуального розвитку.
Регулювання гормональної активності більшості залоз внутрішньої секреції здійснюється за принципом від'ємного зворотного зв'язку: сам гормон (його кількість у крові) регулює своє утворення.
Зазначений вплив опосередковується через утворення відповідних рилізинг-гормонів. Так, наприклад, за умови підвищення у крові рівня гормону кори надниркових залоз - кортизолу в гіпоталамусі вивільняється менше АКТГ-РГ, у результаті чого секреція гіпофізом АКТГ знижується. Це призводить до зменшення утворення кортикостероїдів наднирковими залозами і до зниження вмісту кортизолу в крові. Крім того, зворотний зв'язок у цій системі регулювання може бути опосередкований безпосередньо самими гормонами гіпоталамуса й аденогіпофіза.
Наприклад, утворення кортизолу корою надниркових залоз різко збільшується під час стрес-реакцій. Цьому передує збільшення секреції АКТГ-РГ і вихід АКТГ.
Центральне регулювання гіпоталамо-гіпофізарної системи здійснюється центрами, що розташовані в передзоровому полі, у лімбічній системі та структурах стовбура мозку (довгастому і середньому мозку, мості). Сигнали від цих центрів до ядер гіпоталамуса передаються за допомогою нейронів, що належать до моноамінергічних систем головного мозку, трансмітерами в яких є біогенні аміни (норадреналін, дофамін, серотонін).
У ЦНС утворюються нейроендокринні пептиди (нейрогормони)-ендорфіни, енкефалін.
Ендорфін має анальгезувальну дію, енкефаліни відіграють роль нейротрансмітерів.
Найбільш важливими гіпоталамічними нейротрансмітерами є дофамін, норадреналін, адреналін.
До зазначених центрів ЦНС надходять не тільки нервові імпульси, але й інформація про рівень гормонів у крові завдяки наявності на мембрані нейронів зазначених відділів рецепторів, сприйнятливих до різних гормонів. Через моноамінові системи гормони впливають на специфічні структури гіпоталамуса і на продукцію ними РГ.
Гормони виділяються для забезпечення того або іншого регуляторного акту, тобто того або іншого функціонального стану організму. І вони так само, як і автономна нервова система, виявляють трофотропну й ерготропну дії.
Трофотропна регуляція - це участь гормонів у регуляції процесів росту й розвитку організму з метою забезпечення його лінійного росту та процесів диференціювання.
Ерготрофна – це участь в регуляції обмінних процесів в організмі.

4.Щитовидна і прищитоподібна залози.
У новонародженого щитоподібна залоза важить 1-2 г (0,16 % маси тіла), 25-30 г — у дорослих (0,05 % маси тіла).
Гормон щитоподібної залози — тиреоглобулін, активною частиною його є тироксин і три-йодтиронін. Півперіод «життя» у плазмі крові тироксину становить 67 діб, трийодтироніну — 2-3 доби.
Механізм дії гормону зводиться до посилювання окислювальних процесів у мітохондріях клітини (зменшення проникності мембран мітохондрій, набухання тощо, роз'єднування процесів фосфорилування і перенесення електронів).
Гіпофункція щитоподібної залози супроводжується мікседемою (слизовий набряк: набухання тканин, збільшення в них колоїду, слизу). Гіпофункція щитоподібної залози, яка з'являється з народження, супроводжується кретинізмом — низький зріст, психічна неповноцінність, затримка статевого дозрівання тощо. Нестача виділення тиреоглобуліну може виникнути в зв'язку зі зниженням вмісту йоду в їжі — ендемічний зоб. Розростання тканини щитоподібної залози є «компенсаторним».
Підвищення функції щитоподібної залози (хвороба Базедова і Гревса) також супроводжується збільшенням тканини щитоподібної залози, але при цьому розростається функціонуюча тканина залози, а не втрачається вміст колоїду в залозі, як при ендемічному зобі.
Надлишок тироксину знижує величину потенціалу спокою гладком'язових клітин і міокардіоцитів, блокує автоматію перших і підвищує чутливість міометрія матки до окситоцину, норадреналіну, що у разі вагітності веде до скорочення матки і до викидня.
Артеріальний тиск при гіпертиреозі підвищений за рахунок тахікардії. Потенціал дрібних і середніх кровоносних судин знижений, вони не пульсують, гіпотонічні, компенсаторно посилюється серцевий викид. Тахікардія при гіпертиреозі зумовлена зниженням мембранного потенціалу провідної системи серця
Ендокринна функція прищитовидних залоз
У людини є дві пари прищитовидних залоз, які розташовані на поверхні або в товщі щитовидної залози. В окремих випадках загальна кількість цих залоз у людини може складати 6 або 8. Прищитовидні залози виробляють паратгормон, який разом із тирекальцитоніном регулює вміст іонів Са в крові. Основні фізіологічні ефекти паратгормону протилежні до ефектів тиреокальцитоніну. Він збільшує концентрацію Са+2 в крові. Це досягається за рахунок: резорбції Са+2 із кісток, яка зв'язана в активацією остекластів; посилення реабсорбції Са+2 в дистальних ниркових канальцях; посилення всмоктування Са+2 в шлунково-кишковому тракті під впливом продукту перетворення вітаміну Д3 в нирках - кальцитріолу. Одночасно паратгормон стимулює звільнення фосфатів з мінеральної речовини кісток і гальмує реабсорбцію фосфатів в нирках, знижуючи їх концентрацію в крові.
Читайте також:
- I. Особливості аферентних і еферентних шляхів вегетативного і соматичного відділів нервової системи
- IV. Розподіл нервової системи
- IV. Система зв’язків всередині центральної нервової системи
- VI. Філогенез нервової системи
- Адаптація нервової системи до фізичних навантажень
- Адмін і судове оскарження – відмінності.
- Анатомічні особливості вегетативної нервової системи.
- АНАТОМІЯ І ФІЗІОЛОГІЯ ЦЕНТРАЛЬНОЇ ТА ПЕРИФЕРИЧНОЇ НЕРВОВОЇ СИСТЕМИ, ЇЇ ВІКОВІ ОСОБЛИВОСТІ
- Анатомо-фізіологічні особливості нервової системи у новонароджених
- Біологічна регуляція, її види і значення. Контур біологічної регуляції. Роль зворотнього зв’язку в регуляції.
- Біологічне значення нервової системи
- Біологічне значення нервової системи
| <== попередня сторінка | | | наступна сторінка ==> |
| | | Механізм дії паратгормона зв'язаний з аденілатциклазною системою. |
|
Не знайшли потрібну інформацію? Скористайтесь пошуком google: |
© studopedia.com.ua При використанні або копіюванні матеріалів пряме посилання на сайт обов'язкове. |