РЕЗОЛЮЦІЯ: Громадського обговорення навчальної програми статевого виховання
ЧОМУ ФОНД ОЛЕНИ ПІНЧУК І МОЗ УКРАЇНИ ПРОПАГУЮТЬ "СЕКСУАЛЬНІ УРОКИ"
ЕКЗИСТЕНЦІЙНО-ПСИХОЛОГІЧНІ ОСНОВИ ПОРУШЕННЯ СТАТЕВОЇ ІДЕНТИЧНОСТІ ПІДЛІТКІВ
Батьківський, громадянський рух в Україні закликає МОН зупинити тотальну сексуалізацію дітей і підлітків
Відкрите звернення Міністру освіти й науки України - Гриневич Лілії Михайлівні
Представництво українського жіноцтва в ООН: низький рівень культури спілкування в соціальних мережах
Гендерна антидискримінаційна експертиза може зробити нас моральними рабами
ЛІВИЙ МАРКСИЗМ У НОВИХ ПІДРУЧНИКАХ ДЛЯ ШКОЛЯРІВ
ВІДКРИТА ЗАЯВА на підтримку позиції Ганни Турчинової та права кожної людини на свободу думки, світогляду та вираження поглядів
- Гідрологія і Гідрометрія
- Господарське право
- Економіка будівництва
- Економіка природокористування
- Економічна теорія
- Земельне право
- Історія України
- Кримінально виконавче право
- Медична радіологія
- Методи аналізу
- Міжнародне приватне право
- Міжнародний маркетинг
- Основи екології
- Предмет Політологія
- Соціальне страхування
- Технічні засоби організації дорожнього руху
- Товарознавство продовольчих товарів
Тлумачний словник
Авто
Автоматизація
Архітектура
Астрономія
Аудит
Біологія
Будівництво
Бухгалтерія
Винахідництво
Виробництво
Військова справа
Генетика
Географія
Геологія
Господарство
Держава
Дім
Екологія
Економетрика
Економіка
Електроніка
Журналістика та ЗМІ
Зв'язок
Іноземні мови
Інформатика
Історія
Комп'ютери
Креслення
Кулінарія
Культура
Лексикологія
Література
Логіка
Маркетинг
Математика
Машинобудування
Медицина
Менеджмент
Метали і Зварювання
Механіка
Мистецтво
Музика
Населення
Освіта
Охорона безпеки життя
Охорона Праці
Педагогіка
Політика
Право
Програмування
Промисловість
Психологія
Радіо
Регилия
Соціологія
Спорт
Стандартизація
Технології
Торгівля
Туризм
Фізика
Фізіологія
Філософія
Фінанси
Хімія
Юриспунденкция
Обмін нуклеїнових кислот
Hyклеїнові кислоти відіграють основну роль у збереженні та реалізації генетичної інформації. Розрізняють два типи нуклеїнових кислот: дезоксирибонуклеїнові кислоти[ДНК (DNA)], які забезпечують збереження інформації, та рибонуклеїнові кислоти [PHK (RNA)], які беруть участь у процесах генної експресії та біосинтезу білка.
Нуклеїнові кислоти побудовані із нуклеотидних залишків, які в свою чергу складаються з азотистої основи, вуглецевого залишку та фосфатної групи. ДНК і РНК розрізняються за типом вуглецевого залишку та структурою основ.
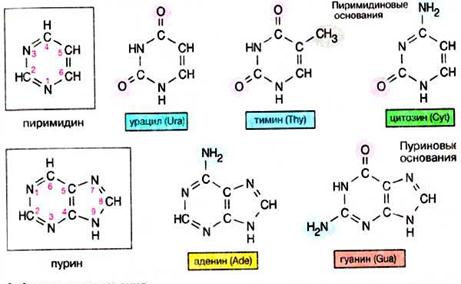
| Азотисті основи —це ароматичні гетероциклічні сполуки, похідніпиримідинуабо пурину. П’ять сполук цього класу є основними структурними компонентами нуклеїнових кислот (загальними для всієї живої матерії). Пуринові основи аденін(Ade) та гуанін(Guа), а також пиримідинова основа цитозин(Cyt), входять до складу ДНК і РНК. До складу ДНК входить також тимін (Thy), 5-метил-похідна урацилу. Основа урацил(Ura) входить тільки до складу РНК. В ДНК вищих організмів в незначній кількості присутній 5-метилцитозин. Урацил за участю ферментів дегідрогенази дигідроурацилу, гідрази дигідропіримідинів і N-карбаміл-β-аланінамідогідролази перетворюється в β-аланін, і карбамінову кислоту (NH2СООН), яка розпадається нааміак і вуглекислий газ. Біосинтез рибонуклеотидів і дезоксирибонуклеотидів є життєво важливим , оскільки ці біомолекули є прямими попередниками ДНК, РНК та нуклеотидних коферментів. Утворення нуклеотидів в організмі відбувається шляхом синтезу з простих попередників, а аткож з готових пуринових і піримідинових основ. |
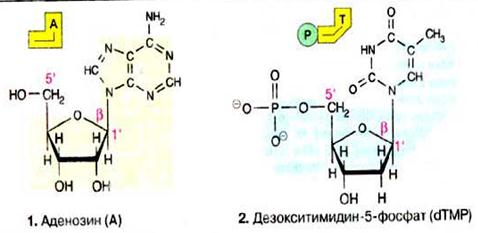
| Сполуки азотистих основ з рибозою або похідні інших азотистих сполук носять назви гуанозин (G), уридин (U), тимідин (T) та цитидин (С). Якщо вуглецевий залишок представлений 2-дезоксирибозою, то утворюється дезоксинуклеозид. В клітині 5'-ОН-группа вуглецевого залишку нуклеозиду етерифікована фосфорною кислотою. Відповідна похідна 2'-дезокситимидину (dT), звено ДНК, називається 2'-дезокситимидин-5'-монофосфат(dTMP) (2). Якщо 5'-фосфатний залишок сполучається з другими нуклеозидфосфатними залишками, одержують нуклеозиди ‑ та нуклеозидтрифосфати (АДФ і АТФ) — важливі коферменти енергообміну. Всі нуклеозидфосфати об’єднані загальною назвою‑ нуклеотиди. В нуклеозидах і нуклеотидах пентоза знаходиться у фуранозній формі. Вуглецевий залишок і азотиста основа сполучені N-глікозидним зв’язкомміж С-1' вуглецевого залишку і N-9 пуринового або відповідно N-1 піримідинового циклу. Глікозидний зв’язок знаходиться в β-конфігурації.. |
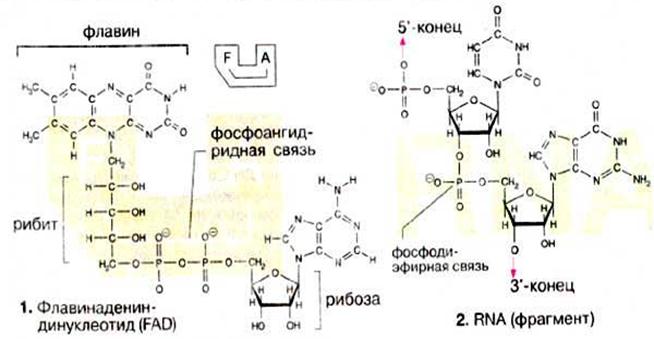
Залишки фосфорної кислоти могуть сполучатися за рахунок утворення фосфоангідридного зв’язку. Отже, два нуклеотиди можуть бути сполучені через фосфатні групи з утворенням відповідного динуклеотиду. До цієї групи сполук належать коферменти [HAДФ+(NADP+)] і KoA(CoA), а також флавін [ФАД(FAD)].
Такий динуклеотид несе на 5'-кінці вільну фосфатну групу, а на 3'-кінці вільну ОН-групу. Тому можна за рахунок утворення ще однієї фосфодиефірного звязку приєднати новий мононуклеотид. Так утворюються олігонуклеотидиі полінуклеотиди.
Полінуклеотиди, утворені з рибонуклеотидних залишків, називаються рибонуклеїновими кислотами (РНК), з дезоксирибонуклеотидних мономерів — дезоксирибонуклеїновими кислотами(ДНК). При позначенні полінуклеотидів вказивають скорочені назви нуклеозидних фрагментів у напрямі 5'→3', (зліва направо). Іноді до назви включають фосфатну групу ("p"). Наприклад, фрагмент РНК, наведений на схемі 2, можна записати ...pUpG... або скорочено ..UG...
|

 Гідроліз нуклеїнових кислот
Гідроліз нуклеїнових кислот

|
 |
8  НуклеопротеїдиНСІпротеїни (протаміни, гістони) + нуклеїнові кислоти
НуклеопротеїдиНСІпротеїни (протаміни, гістони) + нуклеїнові кислоти
8  Нуклеїнові кислотинуклеази олігонуклеотиди → мононуклеотиди
Нуклеїнові кислотинуклеази олігонуклеотиди → мононуклеотиди
8 
 Нуклеотидикомплекс ферментів пуринові та піримідинові основи + рибоза та дезоксирибоза + фосфорна кислота
Нуклеотидикомплекс ферментів пуринові та піримідинові основи + рибоза та дезоксирибоза + фосфорна кислота
Нуклеотиди і нуклеозиди всмоктуються з кишок у кров і доставляються у клітини організму.
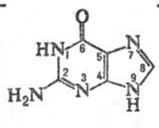
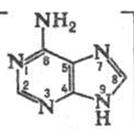
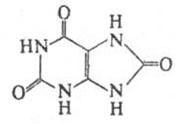
Похідні, які містять систему пурину, широко поширені в природі та відіграють значну роль в багатьох біологічних процесах. Найважливіші похідні пурину ‑ аденін, гуанін (пуринові основи), гіпоксантин, кофеїн, сечова кислота. Ядро пурину входить до складу деяких антибіотиків і нуклеотидів , які є структурними фрагментами нуклеїнових кислот. Пурин і ряд його похідних виявляють протипухлинну, противірусну та протиалергічну активність.
Кінцевим продуктом обміну пуринових основ у клітинах організму є сечова кислота, наприклад у випадку аденіну. При захворюванні людини подагрою сечова кислота відкладається в суглобах, викликаючи при рухах нестерпний біль. Виводиться з сечею. Утворює кислі солі – урати. При деяких захворюваннях вони накопичуються в органах і тканинах.

| 
| 
| 
| 
|
| пурин |  гуанін гуанін
| аденін | гіпоксантин | сечова кислота |
| Пуринові основи |

| Кофеїн (1,3,7- триметилксантин) – пуриновий алкалоїд міститься в бобах кави, горіхах кола, листі чаю (після ферментації). Біосинтез кофеїну в рослинах відбувається через утворення ксантину (утворюється з аденіну і гуаніну). Кофеїн збуджує центральну нервову систему, звужує кровоносні судини, підвищує кров’яний тиск і стимулює серцеву діяльність, стимулює діурез. Використовується при інфекційних та інших захворюваннях, що супроводжуються пригніченням функцій центральної нервової і серцево-судинної систем, при отруєннях наркотиками, при спазмах судин головного мозку, для підвищення психічної та фізичної активності. Кофеїн входить до складу напою "кола". |
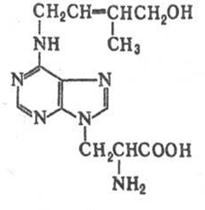
| Теобромін (3,7 – диметилксантин) ‑ безбарвні кристали, мало розчинний у воді (1:700), етанолі, діетиловому ефірі, розчиняється у водних розчинах лугів і кислот. Міститься в какао (1,5-2%) та парагвайському чаї (Ilex paraquariensis). Теобромін спричиняє стимулюючу дію на серцеву діяльність, розширює судини серця і мускулатуру бронхів, підвищує сечовиділення. Використовують при спазмах судин мозку, рідко ‑ при набряках серцевої та ниркової етіології. Одержують сублімацією з бобів какао і листя чаю. |
Обмін речовин в організмі проходить як єдине ціле при тісній взаємодії і взаємній зумовленості окремих його процесів. Навіть перший етап – є одночасний процес розщеплення в травному тракті білків, ліпідів і вуглеводів. Подальші перетворення білків, ліпідів і вуглеводів у тканинах в процесах обміну тісно пов’язані між собою.
При порушенні обміну одного з трьох класів органічних сполук в організмі виникають паталогічні зміни, які впливають на обмін інших речовин. З біохімічної точки взаємозв’язок між перетвореннями вуглеводів, білків і ліпідів виявляється в тому, що з вуглеводів їжі за певних умов можуть синтезуватися ліпіди, а з білків – вуглеводи. Тому при нестачі вуглеводів потреба в білках зростає, а недостане надходження ліпідів може компенсуватись вуглеводами.
1. Взаємозв’язок між обміном вуглеводів і ліпідів:
Ø Синтез жирів з вуглеводів.
Гліцерин утворюється як проміжний продукт анаеробного розщеплення вуглеводів, перетворюється у фосфорогліцериновий альдегід, який легко перетворюється на піровиноградну кислоту (центральний метаболіт вуглеводного обміну). Піровиноградна кислота в результаті окислювального декарбоксилювання і утворення ацетил-КоА перетворюється до ацетооцтової кислоти, з якої далі синтезується одноосновні високомолекулярні кетокислоти, що беруть участь в утворення жирних кислот. Активна форма гліцерину і жирні кислоти використовуються для синтезу жиру.
Ø Синтез вуглеводів з ліпідів.
В результаті гідролізу ліпідів утворюються гліцерин і жирні кислоти. Під час окислення жирних кислот і окислювального декарбоксилювання піровиноградної кислоти утворюється ацетил-КоА, який є сполучною ланкою між обміном вуглеводів і ліпідів. Але не весь ацетил – КоА, що утворився під час окислення жирних кислот, використовується для сине туз вуглводів. Пояснення: можливість депонування вуглеводів в організмі обмежена порівняно з жирами, які є основним енергетичним матеріалом. ЕЕнергетична цінність жирів приблизно у 2,5 рази вища, ніж вуглеводів. Тому відкладання жиру дає змогу тваринам створювати рухомий резерв енергетичного матерівіалу, необхідного для забезпечення перебігу метаболічних реакцій.
Крім синтезу вуглеводів з ацетил-КоА незначна кількість їх може утворюватись з гліцерину шляхом окислення до гліцеринальдегіддегідромонофосфата, який здатний окислюватися до ацетил-КоА.
2. Взаємозв’язок між обміном вуглеводів і білків:
Ø Синтез вуглеводів з білків (глюконеогенез).
Процес перетворення білків на вуглеводи за звичайних фізіологічних умов відбувається незначною мірою, оскільки розщеплення амінокислот направлене переважно в бік утворення кінцевих продуктів. Важливе значення глюконеогенез має в екстремальних умовах, за яких необхідна адаптація організму до зміни умов зовнішнього середовища, коли енерговитрати організму збільшуються і значно підвищуються витрати глікогену.
Найінтенсивніше вуглеводи синтезуються з глюкогенних амінокислот (аспарагінова, глутамінова кислоти, валін, серин, орнітін, цистеїн, гліцин, аланін), які після переамінування і дезамінування перетворюються на кетокислоти: щавлевооцтова, α-кетоглутарова, піровиноградна. ПВК – основний метаболіт, який забезпечує перетворення амінокислот на вуглеводи. Щавлевооцтова та α-кетоглутарова кислоти після декарбоксилювання також перетворюються на піровиноградну кислоту. Перетворення ПВК у вуглеводи може здійснюватися через ацетил-КоА, який утворюється під час ї окислювального декарбоксилювання або внаслідок протилежно напрямлених реакцій гідролізу.
Ø Синтез білків з вуглеводів.
З вуглеводів в організмі утворюються піровиноградна, щавлевооцтова, кетоглутарова кислоти, які після амінування перетворюються на відповідні амінокислоти
З глутамінової кислоти можуть синтезуватися пролін, гідроксипролін, орні тин.
Отже, з метаболітів вуглеводного обміну в організмі можуть синтезуватися більшість амінокислот, крім незамінних. Цим пояснюється незамінність білків у харчуванні. Процес синтезу амінокислот з вуглеводів має обмежене значення.
3. Взаємозв’язок між обміном білків і ліпідів:
Ø Утворення ліпідів з білків.
Експериментально встановлено, що при значному вмісті білків у харчовому раціоні частина їх перетворюється на жири. Сполучною ланкою у цьому процесі є піровиноградна кислота, яка утворюється під час дезамінування і пере амінування аланініу, серину та інших амінокислот.
ПВК внаслідок декарбоксилювання перетворюється на Ацетил–КоА, який може використовуватися для синтезу вищих жирних кислот.
Амінокислоти, які в процесі перетворення утворюють метаболіти, можуть використовуватись для синтезу жирів, називаються кетогенними.
З продуктів білкового обміну може синтезуватись також гліцерин. Аланін легко перетворюється в ПВК, яка під час зворотного гліколізу перетворюється в діоксиацетонмонофосфат – спільний метаболіт вуглеводного і жирового обміну, а далі легко перетворюється на гліцерин. Частина амінокислот використовується для синтезу азотовмісної частини складних ліпідів (фосфатидів).
Ø Утворення білків з ліпідів
Цей процес в організмах людини і тварин має обмежене значення. Частковий синтез замінних амінокислот з жирів може здійснюватись з кетокислот – піровиноградної, α-кетоглутарової, щавлевооцтової кислот, які легко піддаються амінуванню і переамінуванню. Оскільки з жирів утворюється незначна кількість кетокислот, тому жири не можуть бути важливим джерелом синтезу білків.
Висновок:
1 Процеси обміну білків, ліпідів, вуглеводів в організмі взаємопов’язані.
1 Спільним метаболітом, який використовується в процесах взаємоперетворення, є піровиноградна кислота.
1 Піровиноградна кислота перетворюється в ацетил-КоА, який перетворюється в жирні кислоти, які під час взаємодії з гліцерином утворюють жири.
1 Внаслідок карбоксилювання піровиноградної кислоти утворюється щавлевооцтова кислота, а далі вона може утворювати дикарбонові амінокислоти – аспарагінову і глутамінову. Ці амінокислоти в реакціях переамінування сприяють біосинтезу всіх інших замінних амінокислот.
1 Піровиноградна кислота здатна відновлюватись до молочної кислоти, яка використовується в процесах синтезу вуглеводів у тканинах організму.
1 Оцтова кислота виступає у вигляді активованої форми – ацетилкоферменту А (ацетил-КоА), який виконує роль центральної проміжної ланки, через яку відбуваються процеси окислювального розпаду різних сполук і з стадії якої беруть початок численні синтетичні реакції в клітині.
1 Цикл трикарбонових кислот (ЦТК) є тією стадією, на якій сходяться всі види обміну речовин, втрачаючи свою специфіку. Саме в цьому циклі відбувається повне об’єднання обміну білків, вуглеводів і ліпідів у єдиний процес і саме звідси починаються взаємні перетворення цих речовин.
Схема перетворення вуглеводів на ліпіди
| Вуглеводи | ||||||||

| ||||||||
 Глюкоза Глюкоза
| Гліколіз | 
|   Гліцеринальдегідофосфат Гліцеринальдегідофосфат
| 
| Фосфогліцеринова кислота
| |||
| Ліпіди | Діоксіацетонмонофосфат | Піровиноградна кислота | ||||||
 
|
|
| ||||||
 Гліцерин + Вищі жирні кислоти Гліцерин + Вищі жирні кислоти
| Гліцерофосфат | Ацетил-КоА | ||||||
 
|
|  
| ||||||

| ||||||||
| ЦТК |
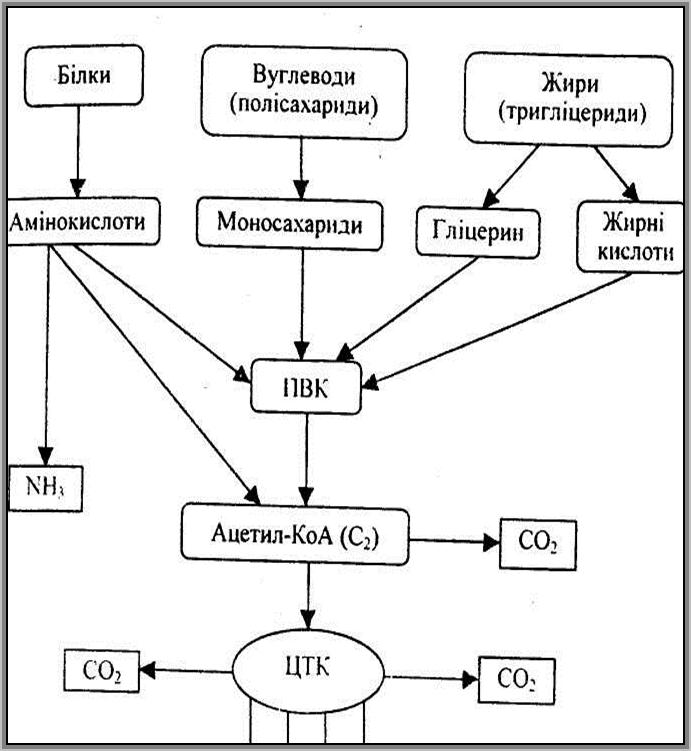
Схема взаємоперетворення білків, вуглеводів і ліпідів
 Вуглеводи Вуглеводи
| Жири | Білки | ||
| 
| |||
| Гліколіз | Гідроліз | |||
| β-окислення жирних кислот | 
| ||
| Піровиноградна кислота | 
| Амінокислоти | ||

|
| |||
| Окислювальне декарбоксилювання | 
| β-кетокислоти | ||
  Ацетилхолін Ацетилхолін
|  Синтез жирних кислот Синтез жирних кислот
| Обмін амінокислот | ||

| ||||

| Ацетил-КоА | |||
| Стероїди | 
| |||
| Цикл Трикарбонових кислот | ||||

| ||||
| СО2 + Н2О | ||||
Читайте також:
- Автоматичний обмін даними.
- АДАПТАЦІЯ ОБМІНУ РЕЧОВИН ДО М'ЯЗОВОЇ ДІЯЛЬНОСТІ
- Альдегід Реактив Толенса Кислота
- Американський стандартний код для обміну інформацією ASCII.
- Білки – це високомолекулярні органічні біополімери, мономерами яких є амінокислоти.
- Білковий обмін
- Білковий обмін: загальні відомості
- БІОСИНТЕЗ АМІНОКИСЛОТ
- БІОСИНТЕЗ ЖИРНИХ КИСЛОТ
- Біотехнологічне виробництво незамінних амінокислот
- Буфер обміну — це тимчасове місце зберігання інформації, яку було скопійовано або переміщено з одного місця з метою використання в іншому.
- Вентиляція. Види вентиляції. Організація повітрообміну в приміщеннях, повітряний баланс, кратність повітрообміну
| <== попередня сторінка | | | наступна сторінка ==> |
| Метаболізм жирів | | | Особливості типології у країнознавстві |
|
Не знайшли потрібну інформацію? Скористайтесь пошуком google: |
© studopedia.com.ua При використанні або копіюванні матеріалів пряме посилання на сайт обов'язкове. |