РЕЗОЛЮЦІЯ: Громадського обговорення навчальної програми статевого виховання
ЧОМУ ФОНД ОЛЕНИ ПІНЧУК І МОЗ УКРАЇНИ ПРОПАГУЮТЬ "СЕКСУАЛЬНІ УРОКИ"
ЕКЗИСТЕНЦІЙНО-ПСИХОЛОГІЧНІ ОСНОВИ ПОРУШЕННЯ СТАТЕВОЇ ІДЕНТИЧНОСТІ ПІДЛІТКІВ
Батьківський, громадянський рух в Україні закликає МОН зупинити тотальну сексуалізацію дітей і підлітків
Відкрите звернення Міністру освіти й науки України - Гриневич Лілії Михайлівні
Представництво українського жіноцтва в ООН: низький рівень культури спілкування в соціальних мережах
Гендерна антидискримінаційна експертиза може зробити нас моральними рабами
ЛІВИЙ МАРКСИЗМ У НОВИХ ПІДРУЧНИКАХ ДЛЯ ШКОЛЯРІВ
ВІДКРИТА ЗАЯВА на підтримку позиції Ганни Турчинової та права кожної людини на свободу думки, світогляду та вираження поглядів
- Гідрологія і Гідрометрія
- Господарське право
- Економіка будівництва
- Економіка природокористування
- Економічна теорія
- Земельне право
- Історія України
- Кримінально виконавче право
- Медична радіологія
- Методи аналізу
- Міжнародне приватне право
- Міжнародний маркетинг
- Основи екології
- Предмет Політологія
- Соціальне страхування
- Технічні засоби організації дорожнього руху
- Товарознавство продовольчих товарів
Тлумачний словник
Авто
Автоматизація
Архітектура
Астрономія
Аудит
Біологія
Будівництво
Бухгалтерія
Винахідництво
Виробництво
Військова справа
Генетика
Географія
Геологія
Господарство
Держава
Дім
Екологія
Економетрика
Економіка
Електроніка
Журналістика та ЗМІ
Зв'язок
Іноземні мови
Інформатика
Історія
Комп'ютери
Креслення
Кулінарія
Культура
Лексикологія
Література
Логіка
Маркетинг
Математика
Машинобудування
Медицина
Менеджмент
Метали і Зварювання
Механіка
Мистецтво
Музика
Населення
Освіта
Охорона безпеки життя
Охорона Праці
Педагогіка
Політика
Право
Програмування
Промисловість
Психологія
Радіо
Регилия
Соціологія
Спорт
Стандартизація
Технології
Торгівля
Туризм
Фізика
Фізіологія
Філософія
Фінанси
Хімія
Юриспунденкция
План лекцій
Лекція 4. Обмін речовин мікроорганізмів
Контрольні запитання
1. Яка номенклатура прийнята для визначення назви прокаріотів?
2. Розмір, форма і будова дріжджових клітин.
3. Способи розмноження дріжджів.
4. У чому полягає сутність брунькування дріжджів?
5. Які функції виконують спори у дріжджів на відміну від бактерій?
6. Класифікація дріжджів. Характеристика їх родин та окремих родів і видів.
7. Назвіть представників справжніх дріжджів, які найчастіше використовуються в харчовій промисловості.
8. Які дріжджі найчастіше спричиняють псування харчових продуктів?
9. Будова тіла і клітини плісеневих грибів.
10. Способи розмноження плісеневих грибів.
11. У чому полягає відмінність ендоспор та аскоспор?
12. Чим відрізняються аскоспори від базидіїв?
13. Систематика плісеневих грибів.
14. Назвіть найпоширеніших збудників псування харчових продуктів і промислових товарів серед плісеневих грибів.
15. Що таке віруси? Яку будову вони мають?
16. Які фази взаємодії вірусів та організму-господаря Вам відомі? Охарактеризуйте їх.
1. Хімічний склад мікробної клітини.
2. Ферменти, їх роль у забезпеченні життєдіяльності мікроорганізмів.
3. Живлення мікроорганізмів - конструктивний обмін.
4. Дихання мікроорганізмів - енергетичний обмін.
Обмін речовин і енергії – або метаболізм – це сукупність хімічних і фізичних перетворень речовин і енергії, які відбуваються у живому організмі і забезпечують його життєдіяльність. Обмін речовин і енергії складає єдине ціле і підпорядковується універсальному закону збереженням матерії і енергії. Метаболізм забезпечує гомеостаз організму, забезпечує організм енергією та ряду інших проявів життя. Обмін речовин складається із процесів асиміляції і дисиміляції. Асиміляція (анаболізм) – процес засвоєння організмом речовин, за якого поглинається енергія. Дисиміляція (катаболізм) – це процес розпаду складних органічних сполук, який проходить із виділенням енергії.
Підфізіологією мікроорганізмів розуміють хімічний склад мікробної клітини і різні процеси, пов'язані з її життєдіяльністю. Мікробні клітини майже цілком складаються з води (біля 80 %). Лише 20 % вмісту клітини припадає на сухі речовини. Якщо їх прийняти за 100 %, то хімічний склад клітини буде такий: вуглецю - 46-50%, кисню – 30%, водню - 6-7%, азоту - 7-14%, мінеральних речовин - 2-14%. До мінеральних речовин належать фосфор, калій, натрій, магній, сірка, кальцій, хлор, залізо, цинк, бор, хром та інші. Усе необхідне клітина одержує разом з білками, жирами, вуглеводами та іншими речовинами.
Білки- високомолекулярні органічні сполуки із залишків амінокислот. Вони відіграють важливу роль у життєдіяльності клітин, виконуючи пластичну,
спадкову, каталітичну функції.
Вуглеводи.Розрізняють прості вуглеводи - моносахариди, або сахара (глюкоза, фруктоза) та складні - полісахариди, які поділяють на низькомолекулярні - дисахариди (сахароза, лактоза) і високомолекулярні (крохмаль, клітковина). Вуглеводи є основним енергетичним матеріалом. Завдяки їм у клітині здійснюються процеси утворення та перетворення енергії.
Жири- органічні біополімери, які складаються із гліцерину та жирних кислот. Вони виконують запасну та енергетичну функції у клітині.
Мінеральні речовинипотрібні у невеликій кількості, але за їх відсутності виникають різні зміни у життєдіяльності клітин. Їх умовно можна поділити на структурні, метаболічні та каталітичні. Вони забезпечують ряд життєво важливих процесів. Це стосується насамперед підтримання певних фізико-хімічних констант - осмотичного тиску, кислотно-лужної рівноваги, процесів збудження та гальмування, каталітичної активності ферментів тощо.
Ферменти- специфічні білки, які присутні у живих клітинах і виконують функції біологічних каталізаторів. Завдяки їм реалізується генетична інформація і здійснюються процеси обміну речовин та енергії у живих організмах. Ферменти мають білкову природу і можуть бути простими або складними. Останні містять небілкову частину - кофермент.
1. Особливості ферментів мікроорганізмів, їх функції. Структура, властивості, синтез, функції та класифікація ферментів мікроорганізмів такі ж, як у більш складних організмів. Однієї із особливостей ферментів мікроорганізмів є перевага адаптивних (індуцибельних) ферментів над конститутивними, що зв'язано з малим вмістом протоплазми, роллю головного механізму адаптації до мінливих умов зовнішнього середовища. До особливостей ферментів мікроорганізмів у гетеротрофних мікроорганізмів може бути також віднесене виділення їх у великих кількостях у зовнішнє середовище. Ця група так званих екзоферментів здатна викликати трансформацію (деградування) будь-яких органічних і неорганічних речовин, та включати їх складові у новий цикл круговороту речовин та енергії. Екзоферменти здійснюють контактне і позаклітинне переварювання речовин, беруть участь у процесі самоочищення води та ґрунту від органічних залишків і ін. Визначення ферментів мікроорганізмів використовують у бактеріологічній практиці для ідентифікації. Очищені або у складі бактеріальної клітини ферменти застосовують для одержання великої кількості необхідних для промисловості та медицини препаратів і матеріалів, включаючи мікробний білок, антибіотики, вітаміни й ін. Саме завдяки ферментам здійснюється усі життєвонеобхідні реакції мікроорганізмів
2. Види ферментів та їх роль. Відповідно до типу реакцій, які каталізують ферменти їх розділяють на шість класів:
Оксидоредуктази - каталізують реакції окислення-відновлення.
Трансферази - каталізують реакції переносу різних груп від донора до акцептора.
Гідролази - каталізують розриви зв'язків у субстратах із приєднанням води.
Ліази - каталізують реакції розриву зв'язків у субстраті без приєднання води чи окислення.
Ізомерази - каталізують перетворення в межах однієї молекули (внутрішньомолекулярні перебудови).
Лігази (синтетази) - каталізують приєднання двох молекул із використанням енергії фосфатних зв'язків.
Незважаючи на малі розміри мікробної клітини, розподіл у ній ферментів суворо впорядкований. Ферменти енергетичного обміну і транспорту поживних речовин локалізовані у цитоплазматичній мембрані та її похідних. Ферменти
білкового синтезу зв'язані із рибосомами. Багато ферментів знаходяться у
у цитоплазмі у розчиненому вигляді. Ферменти бактерій підрозділяються на екзо- і ендоферменти.
Ендоферменти функціонують тільки всередині клітини. Вони каталізують реакції біосинтезу й енергетичного обміну. Екзоферменти виділяються клітиною у середовище та каталізують реакції гідролізу складних органічних сполук до більш простих, доступні для асиміляції мікробною клітиною. До них відносять гідролітичні ферменти, які відіграють винятково важливу роль у харчуванні мікроорганізмів. Залежно від умов утворення ферментів їх розділяють на конститутивні та індуцибельні. Конститутивними називають ферменти, синтезовані клітиною незалежно від субстрату, на якому розвиваються бактерії. Наприклад, ферменти гліколізу. Індуцибельні ферменти синтезуються тільки у відповідь на присутність у середовищі необхідного для клітини субстрату-індуктора. Він взаємодіє із репресором, інактивує його, у результаті чого включається генетичний апарат клітини і починається синтез відповідного ферменту. Індукований синтез ферментів відбувається доти, доки в середовищі присутній індуктор. При цьому ферменти синтезуються заново у всіх клітинах одночасно. Індукторами біосинтезу є багато поживних речовин. До індуцибельних відносять більшість гідролітичних ферментів.
Відомі також ферменти, які одержали назву аллостеричних. Крім активного центру у них є регуляторний чи аллостеричний центр, який у молекулі ферменту просторово розділений із активним центром. Аллостеричним (від греч. allos - інший, чужий) він називається тому, що молекули, які зв'язані із цим центром, за будовою (стерично) не схожі на субстрат, але впливають на зв'язування та перетворення субстрату в активному центрі, змінюючи його конфігурацію. Речовини, що зв'язуються із аллостеричним центром, називають аллостеричними ефекторами. Вони впливають через аллостеричний центр на функцію активного центру. Аллостеричні ефектори можуть бути позитивними (активатори) чи негативними (інгібітори). Деякі ферменти, так звані ферменти агресії, руйнують тканини і клітини макроорганізму, обумовлюючи тим самим, поширення патогенних мікроорганізмів і їх токсинів у інфікованих тканинах. До таких ферментів відносять плазмокоагулазу, нейрамінідазу, колагеназу, лецитиназу, гіалуронідазу та деякі інші ферменти. Гіалуронідаза стрептококів, наприклад, розщеплює гіалуронову кислоту в мембранах клітин сполучних тканин макроорганізму, що сприяє поширенню збудників та їх токсинів в організмі, обумовлюючи високу інвазивність цих бактерій. Плазмокоагулаза є головним чинником патогенності стафілококів, тому що бере участь у перетворенні протромбіну в тромбін, що викликає утворення фібриногену. У результаті кожна бактерія покривається плівкою, що охороняє її від фагоцитозу.
3. Використання ферментів мікроорганізмів людиною. Ферменти мікроорганізмів такі, як лігази і рестриктази, знайшли широке застосування в біотехнології, у тому числі в генній інженерії, для одержання різних біологічно активних речовин, продуковані моноклональних антитіл, а також ряду продуктів у легкій та харчовій промисловості.
Ферменти мікроорганізмів характеризують їх біологічні властивості і тому їх досліджують з метою ідентифікації бактерій. Залежно від субстрату гідролітичні ферменти прийнято поділяти на дві великі групи:
- гідролітичні або цукролітичні ферменти, субстратом для яких є різні цукри, а продуктами їх розщеплення - кислоти, спирти, альдегіди, Н2О ;
- протеолітичні ферменти, що розщеплюють білки із утворенням поліпептидів, амінокислот, аміаку, індолу, сірководню.
Для вивчення активності ферментів у процесі ідентифікації мікроорганізмів широко використовують диференційно-діагностичні середовища, до складу яких входять певні субстрати – цукри чи білки. При дослідженні гідролітичної активності бактерій найбільш поширені моносубстратні диференційно-діагностичні середовища Гісса, лактозовмісні середовища Ендо, Левіна, Плоскірєва, дисубстратні середовища Ресселя, полісубстратні середовища Кліглера й Олькеницького. Останні можуть служити і для вивчення протеолітичних властивостей бактерій, тому що процес росту мікроорганізмів супроводжується вивільненням аміаку. Протеолітичні ферменти бактерій визначають також за виділенням індолу, сірководню, розщепленню деяких амінокислот, наприклад, фенілаланіна, лізина, цистина. Протеолітичні ферменти здатні змінювати (розріджувати) желатин, причому, різні види бактерій по - різному змінюють «стовпчик» желатину в пробірці з посівом мікроорганізму. Так, у процесі росту холерного вібріона «стовпчик» желатину приймає форму цвяха, у процесі росту стафілокока - панчохи, синегнійної палички - спостерігається пошарове розрідження середовища.
Окислювально-відновні ферменти, дегідрогенази, каталазу визначають за зміною органічного барвника - акцептора водню. Здатність мікроорганізмів використовувати як джерело вуглецю цитрат оцінюють у спеціальних тестах заснованих на роботі ферментів. У практичних бактеріологічних лабораторіях широко застосовують мікро- та експрес-методи для орієнтовного вивчення біохімічних властивостей мікроорганізмів. Для цієї мети існує безліч тест-систем. Найчастіше використовують систему індикаторних паперів (СІП). СІПи представляють диски фільтрувального папера, просякнені розчинами чи цукрами інших субстратів у сполученні з індикаторами. Такі диски опускають у пробірку з вирощеною культурою. За зміною кольору диска судять про роботу ферменту. Мікротест системи для вивчення ідентифікації ентеробактерій представлені одноразовими пластиковими контейнерами із середовищами, що містять різні субстрати та з додаванням індикаторів. Посів чистої культури мікроорганізмів у такі тест-системи дозволяє швидко виявити здатність бактерій утилізувати цитрати, глюкозу, сахарозу, виділяти аміак, індол, розкладати сечовину, лізин, фенілаланін і т.д.
Дихання мікроорганізмів. Процес надходження до організму кисню, використання його клітинами і тканинами для окислення органічних речовин та виділення із організму вуглекислого газу називається диханням. Процес дихання у бактерій складається із двох етапів – кисневого та без кисневого. Найважливішим на безкисневому етапі енергетичного обміну є розщеплення у клітинах молекул глюкози шляхом гліколізу на дві молекули піровиноградної або молочної кислоти у м’язових клітинах:
С6Н12О6 + 2АДФ + 2Н3РО4 ®2С3Н6О3 + 2АТФ + 2Н2О
У процесі розпаду глюкози беруть участь 13 різних ферментів, фосфорна кислота і АДФ. Під час гліколізу виділяється » 200 кДж енергії. 84 кДж використовується на синтез 2-х молекул АТФ, а решта (116 кДж) виділяється у вигляді теплоти. Організм отримує енергію в умовах дефіциту кисню. Спиртове бродіння – це один із видів перетворення глюкози, коли вона розпадається на 2 молекули етилового спирту (С2Н5ОН) та 2 молекули вуглекислого газу (СО2).
С6Н12О6 + 2Н3РО4 + 2АДФ = 2СО2 + 2С2Н5ОН + 2АТФ + 2Н2О
Процес кисневого розщеплення описується рівнянням:
2С3Н6О3 + 6О2 + 36АДФ + 36Н3РО4 ® 36АТФ + 6СО2 + 42Н2О
При цьому виділяється енергія (2600 кДж) частина якої розсіюється у вигляді тепла (45%), 55% перетворюється в енергію хімічних зв’язків АТФ (1440 кДж).
Сумарне рівняння повногорозщеплення глюкози записується так:
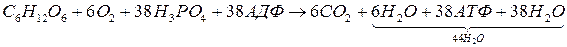
1. Для синтезу АТФ у процесі гліколізу не потрібні мембрани. Він відбувається і в пробірці, якщо є всі ферменти і субстрати.
2. Для кисневого процесу потрібні мітохондріальні мембрани.
3. Розщеплення 1 молекули глюкози до оксиду вуглецю і води забезпечує синтез 38 молекул АТФ. У безкисневій стадії утворюються 2 молекули АТФ, а у кисневій – 36 молекул АТФ).
4. Коли спостерігається нестача кисню, або пошкодження мітохондрій, то клітина, щоб дістати необхідну для життя кількість АТФ використовує безкисневий процес. Для цього їй потрібно в 20 разів більше глюкози, ніж у нормі.
Процес виділення із організму продуктів обміну називають екскрецією. Кінцевими продуктами розщеплення вуглеводів і жирів є СО2 і Н2О, які виводяться з організму. У деяких мікроорганізмів спостерігається виділення невикористаної енергії у формі світлової. Такою здатністю характеризуються деякі бактерії, гриби, протісти. Світіння морської води, дерева, що погнило, риби пояснюється присутністю на них мікроорганізмів, що світяться.
Надходження речовин у клітину та виділення продуктів обміну у довкілля відбувається у мікроорганізмів через всю поверхню тіла. У мікроорганізмів дуже велика поверхня клітини, що обумовлює досить активний обмін речовин. Відомо два шляхи надходження речовин до клітини: осмос і адсорбція (специфічне перенесення). Активна роль у цих процесах належить цитоплазматичній мембрані. Осмос є дифузією речовин у розчинах через напівпроникну перетинку (мембрану). Як відомо, через такі мембрани можуть дифундувати речовини, які знаходяться у стані справжніх розчинів. Осмос виникає під дією різниці осмотичного тиску у розчинах по обидві сторони напівпроникної мембрани. Оболонка клітини проникна і затримує лише макромолекули. Цитоплазматична мембрана клітини характеризується напівпроникністю; вона є осмотичним бар'єром, регулюючи надходження у клітину і вихід із неї розчинених речовин. Речовини, не розчинні у воді або здатні утворювати колоїдні розчини (наприклад, білки, крохмаль), не можуть бути використані клітиною. Вони можуть надходити до неї лише після розщеплювання, поза клітиною, на простіші речовини. Таким чином, при осмотичному надходженні поживних речовин у клітину рушійною силою служить різниця осмотичного тиску між середовищем і клітиною. Якщо осмотичний тиск мікробних клітин вищий ніж у середовищі, то створюється стан тургору у клітинах. При цьому протопласт клітини притискається до клітинної оболонки, злегка розтягуючи її. Якщо мікроорганізми потрапляють у субстрат, осмотичний тиск якого вищий, ніж у клітині, то цитоплазма віддає воду в зовнішнє середовище. Поживні речовини в клітину не надходять, вміст клітини зменшується у об'ємі, і протопласт відстає від клітинної оболонки. Це явище називається плазмолізом клітини. При надмірно низькому осмотичному тиску зовнішнього середовища може настати плазмоптіс клітини — явище, зворотнє плазмолізу, коли внаслідок високої різниці осмотичного тиску цитоплазма швидко переповнюється водою. Це може призвести до розриву клітинної оболонки, що спостерігається, наприклад, при розміщенні бактеріальних клітин у дистильованій воді.
Другий шлях надходження речовин у клітину - активний - шляхом перенесення їх особливими речовинами ферментативної природи. Ці переносники локалізовані у цитоплазматичній мембрані і називають їх пермеазами. Кожен фермент транспортує тільки певну речовину, що має схожу з білком-переносником стереохімічну структуру молекули. На зовнішній стороні цитоплазматичної мембрани переносник адсорбує речовину - вступає з нею у тимчасовий зв'язок та дифундує через мембрану, віддаючи її на внутрішній стороні цитоплазматичної мембрани.
Їжа повинна містити такі речовини, які задовольняли б потребу мікроорганізмів у хімічних елементах, які входять до складу їх тіла. Мікроорганізми відрізняються великою різноманітністю типів живлення. Одні харчуються, подібно до зелених рослин, мінеральними речовинами, синтезуючи із простих речовин складні компоненти клітини. Інші мікроорганізми, подібно до тваринних, потребують органічні сполуки. Вимоги різних мікроорганізмів до поживних речовин, особливо джерел вуглецю і азоту, досить різноманітні та специфічні.
Вуглецеве живлення. Вуглець належить до найважливіших органогенів і складає близько 50% сухої маси клітини. По джерелу вуглецевого живлення мікроорганізми можна розділити на дві групи: автотрофні і гетеротрофні. Автотрофні мікроорганізми використовують як єдине джерело вуглецю для синтезу органічних речовин тіла вуглекислоту та її солі. Синтез органічних речовин із мінеральних сполук вимагає затрати енергії. Серед автотрофних мікроорганізмів є види, які асимілюють вуглекислий газ, як і зелені рослини, використовують сонячну енергію. Їх називають фотосинтезуючими. Інші автотрофні мікроорганізми у процесі синтезу органічних сполук використовують енергію хімічних реакцій окислення деяких мінеральних речовин. Такі мікроорганізми називають хемосинтезуючими. До фотосинтезуючих мікроорганізмів відносять водорості, які мають хлорофіл та деякі пігментні бактерії, наприклад зелені та пурпурні сіркобактерії. У клітинах пурпурних бактерій знаходиться зелений пігмент бактеріохлорофіл, схожий із хлорофілом вищих рослин. У клітинах зелених бактерій також знаходиться у невеликій кількості бактеріохлорофіл, але є і інший фотосинтетичний пігмент, хімічна природа якого не встановлена. Бактеріальний фотосинтез не супроводжується виділенням кисню, як у зелених рослин, а роль води у більшості випадків виконує Н2S, при цьому у клітинах накопичується сірка.
Гетеротрофні мікроорганізми у якості джерела вуглецю використовують органічні сполуки і перетворюють їх у речовини своїх клітин. До таких організмів відносять багаточисленні бактерії, гриби і дріжджі. Більшість гетеротрофних мікроорганізмів живуть за рахунок використання органічних речовин різних субстратів тваринного і рослинного походження. Такі організми називають сапрофітами. Деякі гетеротрофи є паразитами (паратрофами). Вони здатні розвиватися лише у тілі інших організмів, харчуючись їх органічними речовинами. До паразитів належать збудники захворювань людини, тварин і рослин. Багато сапрофітів «всеїдні», тобто здатні, використовувати як джерело вуглецю всілякі органічні сполуки - вуглеводи, спирти, органічні кислоти, білки та ін.
Азотне живлення. Найбільш високими вимогами до джерел азоту відрізняються паразити, які розвиваються лише за рахунок органічних азотовмісних речовин того організму, у якому вони паразитують. Багато сапрофітів (бактерії, гриби, дріжджі) не потребують готових амінокислот, задовольняючись мінеральними сполуками азоту, найкращими із яких є амонійні. Із них та безазотистих органічних речовин ці мікроби синтезують амінокислоти. Джерелом вуглецевого скелета амінокислот найчастіше є різні кетокислоти. Аміак приєднується до кетогрупи і утворює відповідну амінокислоту. Синтезовані амінокислоти, включаючись у реакції переамінування, віддають свої аміногрупи іншим кетокислотам у результаті чого утворюються відповідні амінокислоти. Багато мікроорганізмів (переважно гриби, актиноміцети, рідше бактерії) використовують як джерело азоту нітрати та нітрити. Ці окислені форми азоту заздалегідь відновлюються із утворенням аміаку. Є бактерії і гриби (клас базидіоміцети) здатні використовувати вільний азот атмосфери. Вони переводять його в зв'язаний стан, відновлюючи в аміак, який і використовують для синтезу амінокислот. Ці мікроорганізми називають азотфіксаторами. Амінокислоти, отримані із середовища або синтезовані, служать матеріалом для синтезу білків клітини. Синтез відбувається під направляючою дією ДНК і за участю РНК.
Засвоєння зольних елементів. Для синтезу клітинних речовин потрібні різні зольні елементи: сірка, фосфор, калій, кальцій, магній, залізо. Хоча потреба в них і незначна, проте при дефіциті у поживному середовищі навіть одного із цих елементів мікроорганізми не розвиватимуться і можуть загинути. Більшість мікроорганізмів здатна засвоювати зольні елементи із мінеральних солей. Мікроелементи, необхідні для росту мікроорганізмів, можуть бути використані також у вигляді мінеральних солей. Джерелом кисню і водню є вода та інші речовини.
У складі кожної мікробної клітини є вітаміни, які необхідні для нормальної життєдіяльності. Деякі вітаміни входять до складу простетичних груп ферментів. Одні мікроорганізми повинні отримувати вітаміни у готовому вигляді, інші - добре розвиваються і за відсутності вітамінів у середовищі. Вони здатні самі синтезувати вітаміни із речовин поживного середовища, накопичувати їх у своїх клітинах та виділяти із клітини назовні. Багато мікроорганізмів використовують як своєрідні індикатори для визначення наявності вітамінів у тих або інших продуктах.
Читайте також:
- Види лекцій
- Викладення лекційного матеріалу
- Електронний конспект лекцій
- Завдання лекційних занять
- І. ПРОСЛУХОВУВАННЯ ТА КОНСПЕКТУВАННЯ ЛЕКЦІЙ
- ІІ. МАТЕРІАЛИ ДО КУРСУ ЛЕКЦІЙ
- ІІI. Тематика лекцій
- ІІІ. ПЛАН ПРОВЕДЕННЯ ЛЕКЦІЙНОГО ЗАНЯТТЯ
- Індивідуальний метод обліку кількості знесених яєць проводять у селекційних стадах, застосовують для цього контрольні гнізда, або утримання в індивідуальних клітках.
- КОНСПЕКТ ЛЕКЦІЙ
- Конспект лекцій
- Конспект лекцій
| <== попередня сторінка | | | наступна сторінка ==> |
| Основні методи культивування вірусів | | | План лекції |
|
Не знайшли потрібну інформацію? Скористайтесь пошуком google: |
© studopedia.com.ua При використанні або копіюванні матеріалів пряме посилання на сайт обов'язкове. |