РЕЗОЛЮЦІЯ: Громадського обговорення навчальної програми статевого виховання
ЧОМУ ФОНД ОЛЕНИ ПІНЧУК І МОЗ УКРАЇНИ ПРОПАГУЮТЬ "СЕКСУАЛЬНІ УРОКИ"
ЕКЗИСТЕНЦІЙНО-ПСИХОЛОГІЧНІ ОСНОВИ ПОРУШЕННЯ СТАТЕВОЇ ІДЕНТИЧНОСТІ ПІДЛІТКІВ
Батьківський, громадянський рух в Україні закликає МОН зупинити тотальну сексуалізацію дітей і підлітків
Відкрите звернення Міністру освіти й науки України - Гриневич Лілії Михайлівні
Представництво українського жіноцтва в ООН: низький рівень культури спілкування в соціальних мережах
Гендерна антидискримінаційна експертиза може зробити нас моральними рабами
ЛІВИЙ МАРКСИЗМ У НОВИХ ПІДРУЧНИКАХ ДЛЯ ШКОЛЯРІВ
ВІДКРИТА ЗАЯВА на підтримку позиції Ганни Турчинової та права кожної людини на свободу думки, світогляду та вираження поглядів
- Гідрологія і Гідрометрія
- Господарське право
- Економіка будівництва
- Економіка природокористування
- Економічна теорія
- Земельне право
- Історія України
- Кримінально виконавче право
- Медична радіологія
- Методи аналізу
- Міжнародне приватне право
- Міжнародний маркетинг
- Основи екології
- Предмет Політологія
- Соціальне страхування
- Технічні засоби організації дорожнього руху
- Товарознавство продовольчих товарів
Тлумачний словник
Авто
Автоматизація
Архітектура
Астрономія
Аудит
Біологія
Будівництво
Бухгалтерія
Винахідництво
Виробництво
Військова справа
Генетика
Географія
Геологія
Господарство
Держава
Дім
Екологія
Економетрика
Економіка
Електроніка
Журналістика та ЗМІ
Зв'язок
Іноземні мови
Інформатика
Історія
Комп'ютери
Креслення
Кулінарія
Культура
Лексикологія
Література
Логіка
Маркетинг
Математика
Машинобудування
Медицина
Менеджмент
Метали і Зварювання
Механіка
Мистецтво
Музика
Населення
Освіта
Охорона безпеки життя
Охорона Праці
Педагогіка
Політика
Право
Програмування
Промисловість
Психологія
Радіо
Регилия
Соціологія
Спорт
Стандартизація
Технології
Торгівля
Туризм
Фізика
Фізіологія
Філософія
Фінанси
Хімія
Юриспунденкция
Структура та властивості ферментів
У живих організмах під дією біологічних каталізаторів білкової природи одночасно з величезною швидкістю перебігають тисячі хімічних реакцій, які забезпечують синтез і розпад різноманітних органічних сполук,. Речовини, що виконують біокаталітичні функції, отримали назву "ферменти" – ("fermentum" (лат) - закваска, дріжджі, та "fеrmentatio" – бродіння) і від грецьких слів або "ензими" (enzyme (грец.) - у дріжджах, у заквасці). Термін "фермент" був запропонований на початку ХVІІІ ст. голандським вченим Ван Гельмонтом для речовин, що впливають на спиртове бродіння. У кінці ХVІІ ст. дослідженнями Реомюри та Спаланцані про вплив шлункового соку хижих птахів на м'ясо було показано, що розчинення м'яса - це хімічний, а не механічний процес. У 1836 році Шванн виявив у шлунковому соці фермент пепсин, який перетравлював білки м'яса.
Російський вчений К.С. Кірхгоф вперше показав участь хімічних речовин (ферментів) солоду у перетворенні крохмалю на цукор. На початку ХХ ст. російський фізіолог І.П. Павлов вперше довів, що ферменти можуть існувати в організмі в неактивній формі – у вигляді проферментів. Він показав перетворення проферменту трипсиногену на активний фермент трипсин з участю ентерокінази. У кінці XIX в. німецький вчений Е. Фішер, вивчаючи властивості ферментів, висунув положення, що субстрат підходить до ферменту, як ключ до замка. У 1926 р. Самнер вперше отримав фермент (уреазу) у кристалічному стані і встановив його білкову природу.
На початку ХХ століття з’явилися роботи, присвячені кінетиці ферментативних реакцій. Пізніше були сформульовані теорії механізму дії ферментів, регуляції ферментативної активності. Зараз успішно вивчаються можливості застосування ферментів у медицині, різних галузях науки та промисловості.
Ферменти - високоспеціалізовані універсальні за своєю біологічною функцією білкові сполуки, які синтезуються в процесі життєдіяльності всіх живих організмів і виконують функції каталізаторів біохімічних процесів, тобто прискорюють і здійснюють перебіг певних хімічних реакцій у живому організмі. Ферменти можуть виявляти свою активність як в організмі, так і будучи виділеними з нього.
Значення ферментів для життєдіяльності величезне. У присутності ферментів хімічні процеси в організмі перебігають дуже швидко (інколи за мільярдні долі секунди). Без ферментів ці ж хімічні реакції відбувалися б у мільйони разів довше, що не сумісно з тим рівнем обміну речовин і енергії, який властивий живій матерії. Наприклад, в організмі людини за одну секунду руйнується і синтезується 3 мільйони еритроцитів. До складу одного еритроцита входить 300 -400 мільйонів молекул гемоглобіну – гемпротеїну з молекулярною масою 65 000 – 68 000 Да. Звідси можна уявити собі швидкість хімічних реакцій у живому організмі, які забезпечують ці процеси.
На даний час охарактеризовано декілька тисяч ферментів, понад тисячу з них отримані в хімічно чистому вигляді. Для багатьох білків-ферментів з'ясована їх амінокислотна послідовність.
Вивчення ферментів має величезне значення для будь-якої фундаментальної і прикладної галузі біології, для багатьох практичних галузей хімічної, харчової і фармацевтичної індустрії, зайнятих приготуванням каталізаторів, антибіотиків, вітамінів, амінокислот, пептонів та інших біологічно активних речовин, які використовуються в народному господарстві та медицині. Крім цього, ферменти широко використовуються в науково-дослідних роботах як високоспецифічні реагенти, як інструменти при з'ясуванні будови біополімерів (наприклад, нуклеїнових кислот), при генно-інженерних розробках тощо.
В останні роки вчення про ферменти відокремилося в окрему галузь біохімічної науки – ферментологію (ензимологію), яка розвивається в тісному зв'язку з багатьма науками, зокрема, з органічною, неорганічною та фізичною хіміями, фізіологією, токсикологією, мікробіологією, генетикою, фармакологією тощо. Таким чином, ця галузь знань знаходиться на стику хімічних, біологічних і медичних наук.
1.1.1. Фізико-хімічні властивості білків-ферментів.На сьогоднішній день достеменно відомо, що ферменти – це речовини білкової природи. Підтвердженням цього є факт втрати ферментами бродіння своєї активності під час кип’ятіння, що було досліджено ще Л.Пастером. Кип’ятіння спричинює незворотну денатурацію білка-фермента, внаслідок чого останній втрачає здатність каталізувати хімічну реакцію. Як відомо, білки теж при кип’ятінні денатурують і втрачають сої біологічні властивості. Дія на ферменти різних фізичних і хімічних чинників, таких як вплив УФ- і рентгенівського опромінення, ультразвуку, осадження мінеральними кислотами, лугами, алкалоїдними реактивами, солями тяжких металів тощо теж спричинює денатурацію ферментів, так само як і білків, і втрату їх каталітичної активності. В основі денатурації лежить руйнування зв’язків, що стабілізують вищі структури ферменту-білка (четвертинну, третинну, вторинну) і як наслідок випадання його в осад. Це свідчить про те, що просторова структура білка впливає на виявлення його ферментативної активності.
Аналогічно до білків ферменти під час гідролізу розпадаються на амінокислоти, що, поза сумнівом, підтверджує білкове походження ферментів. У процесі каталізу приймають участь наступні функціональні групи ферментів: СООН-групи дикарбонових амінокислот, NH2-групи лізину, SH-групи цистеїну та дисульфідні цистину, ОН-групи серину та треоніну, гуанідинові групи аргініну, імідазольні групи гістидину, тіоефірні групи метіоніну, фенольні групи тирозину, гідрофобні ланцюги аліфатичних амінокислот і ароматичне кільце фенілаланіну.
Фізико – хімічні властивості перелічених амінокислотних залишків поліпептидного ланцюга ферменту визначають контакт із відповідним субстратом та його перетворення. Гідрофобні радикали амінокислот мають спорідненість до неполярних ділянок субстрату. Полярні групи проявляють кислотні, або основні, або спряжені кислотно-основні (наприклад, гістидин) властивості. Зсув рН середовища викликає зміни їх кислотно-основних властивостей і сприяє контакту з різними групами субстрату.
Водні розчини ферментів є стійкими та гомогенними і можуть тривалий час зберігатися, не випадаючи в осад (не коагулювати), тобто мають властивості справжніх розчинів. Одночасно з тим, за рахунок високої молекулярної маси ферментів їх розчинам притаманні властивості й колоїдних систем.
Доказом білкової природи ферментів слугує виділення їх у чистому вигляді в формі кристалів білка. На сьогоднішній день отримано понад 1 000 кристалічних ферментів. Структура багатьох із них досліджена детально методами рентгеноструктурного аналізу, ядерного магнітного резонансу, електронного парамагнітного резонансу тощо.
Ферменти, як і білки, володіють низкою властивостей, характерних для високомолекулярних сполук: амфотерністю (можуть існувати в розчині в вигляді аніонів, катіонів, амніонів); електрофоретичною рухливістю (завдяки наявності позитивних і негативних зарядів) та втратою рухливості в електричному полі в ізоелектричній точці; вони не здатні до діалізу через напівпроникні мембрани, проте шляхом діалізу їх розчини можна звільнити від низькомолекулярних домішок. Як і білки, ферменти легко осаджуються з водних розчинів методами висолювання чи обережним додаванням ацетону, етанолу та інших речовин, не втрачаючи при цьому своїх каталітичних властивостей.
Заряд молекули ферменту залежить від вмісту в ньому кислих і основних амінокислот. У нативній молекулі ферменту заряди розміщені асиметрично на поверхні білка. Якщо в молекулі ферменту кислі амінокислоти переважають над основними, то його молекула буде мати негативний заряд (поліаніон). І, навпаки, якщо переважають основні амінокислоти, то вона заряджена позитивно, тобто веде себе як полікатіон. В ізоелектричному стані ферменти найменш стабільні і можуть випадати в осад.
Ферменти теж мають велику молекулярну масу, яка може сягати кількох мільйонів. Так, молекулярна маса пепсину становить 32 100 Да, лужної фосфатази – 80 000 Да, лактатдегідрогенази – 140 000 Да, каталази – 248 000 Да, уреази – 480 000 Да, піруватдегідрогеназного комплексу – 4 500 000 Да.
Ферменти чинять високоспецифічний вплив, що теж доводить їх білкову природу. Нарешті, прямим доказом білкової природи ферментів є синтез в лабораторних умовах (лабораторія Б.Мерріфілда, Нью-Йорк, 1969) першого фермента – рибонуклеази, який полягав у послідовному включенні 124 амінокислотних залишків у строгій відповідності з послідовністю амінокислот природного фермента – рибонуклеази підшлункової залози. Штучно синтезований фермент не відрізнявся від природної рибонуклеази ані за хімічними, ані за каталітичними та імунологічними тестами.
Враховуючи білкову природу ферментів, слід зважати на їх стабільність, котра визначається низкою чинників. Так, оптимальною температурою для роботи з ферментами є температура тіла, а для препаративних цілей – використання температури, наближеної до 0◦ С. Слід пам’ятати, що низка ферментів чутлива до зниження температури (мітохондріальний фермент АТФаза, який каталізує розпад АТФ, інактивується при 0◦ С, тоді як при кімнатній температурі залишається стабільним). Для більшості ферментів оптимальним рН середовища є 6,0 – 8,0 (хоча існують винятки). Із препаративною метою часто обезводнюють ферменти (видаляють воду) у вакуумі з замороженого розчину (ліофілізація). Осадження з розчину ферментів спиртом чи ацетоном теж здійснюють при низькій температурі, оскільки при кімнатній температурі ці процедури спричинюють втрату ферментативної активності. Для стабілізації ферментів часто користуються хелатоутворювальними агентами, наприклад, ЕДТА (етилендіамінтетраацетат), який може зв’язувати небажані домішки, які гальмують активність фермента. Однією з обов’язкових умов збереження стабільності фермента є його зберігання в висушеному або замороженому стані, велика кількість ферментів може зберігати свою стабільність у вигляді суспензії в концентрованих розчинах амонію сульфату.
1.1.2. Подібність і відмінність між ферментами і неферментними каталізаторами. Властивості ферментів як біологічних каталізаторів.Ферменти та каталізатори небілкової природи підлягають загальним законам каталізу та мають низку спільних властивостей:
- прискорюють лише енергетично можливі реакції, тобто вони не змінюють константу рівноваги та величину вільної енергії;
- збільшують швидкість хімічної реакції шляхом зниження її енергії активації та, у такий спосіб, наближають реакцію до точки термодинамічної рівноваги;
- не впливають на напрям зворотної реакції, яка визначається співвідношенням концентрацій субстратів і кінцевих продуктів;
- не впливають на положення рівноваги зворотної реакції, а лише пришвидшують її досягнення;
- не входять до складу кінцевих продуктів реакції і виходять з реакції в незміненому вигляді, проте, у низці випадків можуть модифікуватися і навіть розпадатися під впливом кінцевих продуктів реакції (наприклад, цитохром Р-450);
- не витрачаються в процесі каталізу, вивільняючись, вони можуть знову реагувати з новими молекулами субстрату.
Однак, для ферментів характерні і специфічні властивості, які відрізняють їх від інших каталізаторів. Ці відмінності пов'язані з особливостями будови ферментів, які є складними білковими молекулами:
1. ефективність ферментів вища, ніж небілкових каталізаторів: швидкість перебігу реакції за участю ферментів зростає в 108 – 1020 разів (фермент уреаза прискорює гідроліз сечовини в 1014 разів), вони діють у мізерних концентраціях (молекула реніну, який синтезується в шлунку теляти, звурджує за 10 хв при температурі 37ºС 106 молекул казеїногену).
2. Ферменти мають високу специфічність дії, яка обумовлена унікальною структурою активного центра, а також конформаційною та електростатичною комплементарністю між молекулами субстрату та фермента. Кожний фермент каталітично прискорює, зазвичай, одну хімічну реакцію, або ж групу реакцій одного типу. Висока специфічність дозволяє ферментам брати участь у регуляції обміну речовин і направляти його в певному напрямі.
3. Активність ферментів може суттєво змінюватися під впливом сполук, котрі прискорюють (активатори) та сповільнюють (інгібітори) швидкість каталізованої реакції, що дає можливість координувати метаболічні процеси, спрямовані на відтворення живої матерії, підтримання постійності гомеостазу та пристосування до умов середовища.
4. Термолабільність ферментів: ферменти каталізують хімічні реакції при невисокій температурі (37 – 40°С); її зростання призводить до денатурації білкової молекули фермента та, відповідно, зниження швидкості реакції або повної її зупинки. При температурі 100°С майже всі ферменти втрачають свою активність. зниження температури нижче оптимальної тимчасово сповільнює активність фермента внаслідок зменшення швидкості дифузії молекул.
5. залежність активності ферментів від рн середовища: ферменти зазвичай найактивніші в межах вузької концентрації іонів Н+ (фізіологічне значення рН = 6,0 – 8,0). Виключення становлять пепсин (оптимум рН = 2,0) та аргіназа (оптимум рН = 10,0). Зміна рН середовища впливає на стан і ступінь іонізації кислотних і основних груп, які входять до складу фермента в цілому та його активного центра зокрема, що впливає на третинну структуру білка та формування активного фермент-субстратного комплексу.
6. У випадку, коли ферментативний процес являє собою низку послідовних реакцій (метаболічні шляхи) дія ферментів строго впорядкована: продукт однієї ферментативної реакції слугує субстратом для іншої. Їх активність змінюється в залежності від потреб організму в кінцевому продукті.
1.1.3. Структурно-функціональна організація ферментів.Оскільки ферменти – це речовини білкової природи, вони, так само як і білки,можуть бути як простими, так і складними (їх переважна більшість). Для ферментної активності білків важливе значення має збереження їх первинної, вторинної та третинної структур; регуляторним ферментам властива четвертинна структура. Більшість внутрішньоклітинних ферментів є олігомерами, які складаються з декількох протомерів, відносно міцно пов'язаних між собою. Так, глутаматдегідрогеназа (відщеплює два атоми водню від глутамінової кислоти з утворенням α-кетоглутарової кислоти), виділена з печінки бика, складається з 8 великих субодиниць, на які вона може дисоціювати.
Ферменти-прості білки являють собою поліпептидні ланцюги, які при гідролізі розпадаються до амінокислот, їх ще називають однокомпонентними. До них належать пепсин, трипсин, уреаза, рибонуклеаза тощо. Ферменти-складні білки крім поліпептидних ланцюгів містять небілкову частину, їх називають двокомпонентними. Білкову частину двокомпонентних ферментів називають апоферментом (забезпечує специфічність дії та відповідає за вибір типу хімічного перетворення субстрату), а небілкову – кофактором (слугує акцептором і донором хімічних груп, атомів і електронів у каталітичній ділянці активного центра фермента). Молекула складного фермента в цілому називається холоферментом.Зв'язок білкової частини ферменту з небілковою здійснюється за рахунок ковалентних і нековалентних зв’язків і може бути різної міцності. Якщо небілкова частина ферменту міцно пов'язана з білком і в циклі біохімічних реакцій не відділяється від нього, її прийнято називати простетичною групою (наприклад, ФАД, ФМН, біотин тощо). Небілкові компоненти, які слабо пов'язані з білком і легко дисоціюють з комплексу з ферментним білком, називають коферментами (наприклад, НАД+, НАДФ+). Коферменти можуть знаходитися у вільному стані й сполучатися з білковою частиною тільки в момент каталітичної реакції, їх можна розглядати в якості другого субстрату. Один і той самий кофермент може сполучатися з різними апоферментами і брати участь у різних хімічних перетвореннях субстрату (наприклад, піридоксальфосфат може брати участь у реакціях трансамінування чи декарбоксилування). Ферменти, які міцно пов’язані з іонами металів і не втрачають цього зв’язку за умов виділення та фракціонування ферменту, називаються металоферментами. У деяких випадках іони металів не входять до складу ферментів як інтегральні структурні компоненти, а виконують лише функцію їх активаторів (іони Са2+ служать активаторами протеїнкінази С, іони Сl- – a-амілази тощо).
1.1.3.1. Класифікація коферментів.Як було сказано вище,для прояву каталітичної активності більшості ферментів необхідна присутність кофермента. Виключення становлять гідролітичні ферменти (протеази, ліпази, рибонуклеаза). Вони беруть участь в акті каталізу, здійснюють контакт між ферментним білком і субстратом, стабілізують апофермент, який, в свою чергу, посилює каталітичний акт небілкової частини і, крім цього, визначає специфічність ферментів, оскільки одна і та ж за хімізмом небілкова частина може функціонувати в складі різних ферментів.
Хімічна природа коферментів, їх функції в ферментативних реакціях дуже різноманітні. Традиційно до коферментів відносять похідні вітамінів. Проте, окрім них є ще певна кількість небілкових сполук, які беруть участь у прояві каталітичної функції ферментів. За хімічною природою коферменти поділяють на:
– похідні вітамінів (табл.1.1);
– динуклеотиди (похідні нікотинаміду – НАД+, НАДФ+; похідне рибофлавіну – ФАД);
– нуклеотиди – похідні пуринів та піримідинів (АТФ, АДФ, ЦТФ, ЦДФ, УТФ, УДФ);
– комплекси порфіринів з іонами металів;
– S-аденозилметіонін (SAM);
– пептидні (глутатіон).
Таблиця 1.1. Коферменти та відповідні їм вітаміни
| Кофермент | Вітамін-попередник | Функція | Фермент |
| Піридоксаль-фосфат | В6 (піридоксин) | Переамінування, декарбоксилування, рацемізація | Трансамінази, декарбоксилази, рацемази |
| Тіамін-дифосфат | В1 (тіамін) | Окисне декарбоксилування α-кетокислот, перенесення альдегідних групи | Трансальдолаза, транскетолаза |
| Кофермент А | В3 Пантотенова кислота | Перенесення ацильних груп, аеробна деградація та синтез жирних кислот | Ацетилтрансферази Ацилтрансферази |
| Тетрагідро-фолієва кислота | Фолієва кислота | Перенесення С1-груп, біосинтез пуринових нуклеотидів | Формілтрансфераза, тимідилатсинтетаза |
| Біотин | Н Біотин | Реакції карбоксилювання за участю СО2 | Карбоксилаза |
| НАД+, НАДФ+ | РР, Нікотинова кислота,(ніацин) | Зворотне перенесення Н+ та е- | Дегідрогенази (піридинзалежні) |
| ФМН, ФАД | В2 (рибофлавін) | Перенесення Н+ та е- | Дегідрогенази (флавінзалежні) |
| Метил-кобаламін, 5-дезоксиадено-зилкобаламін | В12 (ціанокобаламін) | Перенесення метильних груп, реакції трансметилування, ізомеризації | Метилмалоніл-КоА-мутаза |
| Ліпоєва кислота | N (ліпоєва кислота) | Перенесення ацетильних груп | Піруватдегідрогеназа, кетоглутаратдегідрогеназа |
За типом каталізованої реакції коферменти поділяють на:
 1. Коферменти - переносники атомів водню та електронів (НАД+, НАДФ+, ФМН, ФАД, аскорбінова кислота, кофермент Q, глутатіон, гемінові коферменти (металопорфірини - цитохроми)).
1. Коферменти - переносники атомів водню та електронів (НАД+, НАДФ+, ФМН, ФАД, аскорбінова кислота, кофермент Q, глутатіон, гемінові коферменти (металопорфірини - цитохроми)).
 Похідні вітаміну РР – нікотинамідаденіндинуклеотид (НАД+) та нікотинамідаденіндинуклеотидфосфат (НАДФ+) це окиснені форми коферментів, у яких позитивний заряд несе атом азоту піридинового кільця нікотинаміду:
Похідні вітаміну РР – нікотинамідаденіндинуклеотид (НАД+) та нікотинамідаденіндинуклеотидфосфат (НАДФ+) це окиснені форми коферментів, у яких позитивний заряд несе атом азоту піридинового кільця нікотинаміду:
Як видно зі схеми, субстрат (А) втрачає два атоми водню (2 протони та 2 електрони), але на кофермент переноситься лише 2 електрони і 1 протон, другий протон переходить у середовище. У результаті відновлена форма коферменту втрачає позитивний заряд.
Нікотинаміддинуклеотиди входять до складу багатьох дегідрогеназ, необхідних для синтезу енергії в клітині: вони виступають акцепторами та проміжними переносниками атомів водню на початкових стадіях окиснення вуглеводів, жирних кислот, амінокислот, гліцерину, на стадії циклу Кребса і в термінальних стадіях дегідрування в дихальному та монооксигеназному ланцюгах; вони виступають у ролі алостеричних ефекторів ферментів енергетичного обміну. НАДФН2 як донор атомів водню використовується в біосинтетичних відновних реакціях (синтез жирних кислот, холестерину, гормонів тощо).
Похідні вітаміну В2 – флавінмононуклеотид (ФМН) і флавінаденіндинуклеотид (ФАД) – коферменти, які входять до складу флавінових ферментів, що беруть участь у багатьох окиснювальних реакціях: перенесенні електронів і протонів у дихальному ланцюзі, окисненні пірувату, a-кетоглутарату, жирних кислот, біогенних амінів тощо.
Активною частиною флавінових коферментів є ізоалоксазинова циклічна система, вона може приєднувати два атоми водню (2 електрони та 2 протони):

Флавінові коферменти, на відміну від нікотинамідних, зв’язують два протони.

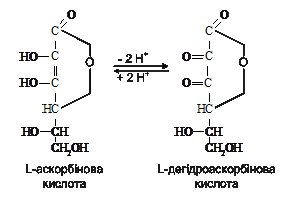 Вітамін С теж бере участь у окисно-відновних процесах. Він може існувати в двох формах – відновленій (аскорбінова кислота) та окисненій (дегідроаскорбінова кислота). Обидві форми легко переходять одна в одну і в якості коферментів гідроксилаз беруть участь в окисно-відновних реакціях:
Вітамін С теж бере участь у окисно-відновних процесах. Він може існувати в двох формах – відновленій (аскорбінова кислота) та окисненій (дегідроаскорбінова кислота). Обидві форми легко переходять одна в одну і в якості коферментів гідроксилаз беруть участь в окисно-відновних реакціях:
Ця властивість обумовлює участь аскорбінової кислоти в обміні білків, вуглеводів, мінеральних речовин.
Металопорфірини за своєю структурою подібні або навіть тотожні гему в гемоглобіні. Порфіринові коферменти входять до таких ферментів як цитохроми (a, b, c), пероксидаза, каталаза тощо.
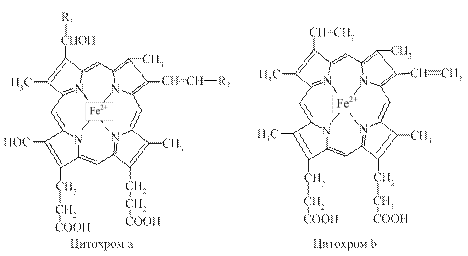
 Вони містять іони металів (зокрема заліза, міді тощо), які здатні змінювати свою валентність (наприклад, Fe2+ « Fe3+) і, у такий спосіб, беруть участь у перенесенні електронів під час окисно-відновних процесів.
Вони містять іони металів (зокрема заліза, міді тощо), які здатні змінювати свою валентність (наприклад, Fe2+ « Fe3+) і, у такий спосіб, беруть участь у перенесенні електронів під час окисно-відновних процесів.
2. Коферменти - переносники різних хімічних груп (нуклеотидні - АТФ, АДФ УТФ, УДФ, ЦТФ, ЦДФ, похідні вітамінів - піридоксальфосфат, піридоксамінфосфат, ліпоєва кислота, КоА, тетрагідрофолієва кислота).
Реакції за участю нуклеотидних коферментів зводяться до перетворення субстрату в молекулі коферменту (наприклад, перетворення УДФ-глюкози на УДФ-галактозу), вони можуть виступати донорами субстратів у реакціях перенесення тих чи інших груп (наприклад, УДФ-глюкоза є донором глюкози в процесі біосинтезу глікогену, ЦДФ-холін – донором холіну в біосинтезі холін фосфатидів тощо).
Піридоксальфосфат і піридоксамінфосфат входять до складу багатьох ферментів у якості коферментів.

Вони відіграють ключову роль в обміні амінокислот: каталізують реакції трансамінування (перенесення аміногрупи з a-амінокислоти на a-кетокислоту) та декарбоксилування (відщеплення карбоксильної групи у вигляді СО2) амінокислот, беруть участь у специфічних реакціях метаболізму окремих амінокислот (серину, треоніну, триптофану, сірковмісних амінокислот, а також у синтезі гему.
Кофермент А (КоА) утворюється з пантотенової кислоти (В3), його сульфгідрильна група може зазнавати ацилування з перетворенням на ацил-КоА, або знаходитися в деацильованому стані (HS-КоА).

Ці коферментні форми беруть участь у перенесенні ацильних радикалів у реакціях загального шляху катаболізму, активуванні жирних кислот, біосинтезі жирних кислот, холестерину, стероїдних гормонів, кетонових тіл, ацетилхоліну, знешкодженні чужорідних тіл у печінці.
Тетрагідрофолієва кислота – відновлена форма ліпоєвої кислоти переносить одновуглецеві залишки (метильні (-СН3), оксиметильні (-СН2ОН), формільні (-НСО) тощо) на різні сполуки і бере тим самим участь у синтезі азотистих основ нуклеїнових кислот, креатину, метіоніну, гліцину, серину.
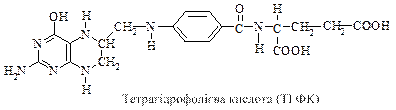
Ліпоєва кислота завдяки своїй здатності легко переходити з окисненої у відновлену форми проявляє властивості коферменту в складі оксидоредуктаз, її амід слугує простетичною групою складного піруват- і a-кетоглутаратдегідрогеназного комплексу, які беруть участь у окисненні піровиноградної та a-кетоглутарової кислот.


3. Коферменти синтезу, ізомеризації та розщеплення вуглець-вуглецевих зв’язків (метилкобаламін, дезоксіаденозилкобаламін, карбоксибіотин, тіаміндифосфат).
Біотин виконує коферментну функцію у якості N5-карбоксибіотину і входить до складу карбоксилаз: він бере участь в утворенні активної форми СО2, використовується в утворенні малоніл-КоА з ацетил-КоА, у синтезі пуринового кільця, у реакції карбоксилування пірувату з утворенням оксалоацетату тощо.
Роль вітаміну В1 визначається тим, що у формі кофактора тіаміндифосфату (ТДФ) він входить до складу як мінімум трьох ферментів і ферментативних комплексів: у складі піруват- і a-кетоглутаратдегідрогеназного комплексів він бере участь в оксиснювальному декарбоксилуванні пірувату та a-кетоглутарату; у складі транскетолази він залучається у пентозофосфатний шлях перетворення вуглеводів.
 Тому цей кофермент необхідний для синтезу жирних кислот, холестерину, стероїдних гормонів, знешкодження токсичних речовин і ліків. Тіамінтрифосфат (ТТФ) у тканині мозку дотичний до синаптичної передачі нервових імпульсів.
Тому цей кофермент необхідний для синтезу жирних кислот, холестерину, стероїдних гормонів, знешкодження токсичних речовин і ліків. Тіамінтрифосфат (ТТФ) у тканині мозку дотичний до синаптичної передачі нервових імпульсів.
 метилкобаламін і дезоксіаденозилкобаламін – це коферментативні форми вітаміну В12. Метилкобаламін тісно пов’язаний із фолієвою кислотою, оскільки входить до складу фермента, який переносить метильну групу 5-метилтетрагідрофолієвої кислоти на гомоцистеїн із утворенням метіоніну.
метилкобаламін і дезоксіаденозилкобаламін – це коферментативні форми вітаміну В12. Метилкобаламін тісно пов’язаний із фолієвою кислотою, оскільки входить до складу фермента, який переносить метильну групу 5-метилтетрагідрофолієвої кислоти на гомоцистеїн із утворенням метіоніну.
Разом із фолієвою кислотою він бере участь у синтезі креатину, азотистих основ, амінокислот, білків, нуклеїнових кислот тощо. Дезоксіаденозилкобаламін бере участь у завершальній стадії окиснення жирних кислот із непарною кількістю вуглецевих атомів, бічного ланцюга холестерину, тиміну, розгалужених амінокислот тощо.
1.1.4. Функціонально активні ділянки ферментів.Біологічна функція як простих, так і складних ферментів обумовлена наявністю в них функціональних ділянок. Під час ферментативної реакції відбувається взаємодія фермента та субстрату (ліганда, який взаємодіє з ферментом) з утворенням проміжних фермент-субстратних комплексів (ФСК). Ділянка молекули фермента, до якої приєднується субстрат, називається активним центром.
 Активний центр простого фермента – це тривимірний утвір, здебільшого щілиноподібної форми, який утворюється сукупністю бічних радикалів амінокислотних залишків, які дуже часто знаходяться на певній відстані в лінійній послідовності поліпептидного ланцюга. Так, у молекулі лізоциму – ферменту, що забезпечує бактерицидні властивості слини, основні групи активного центра представлені амінокислотними залишками, які займають у поліпептидному ланцюзі 35, 52, 62, 63 і 101 положення. У складних ферментах активний центр може містити кофактор, а бічні радикали створюють умови для правильної конформації активного центра, орієнтації та перетворення субстрату. Проте, саме білок у складному ферменті організовує ефективне функціонування кофактора. Так, гем у комплексі з глобіном виявляє свою схильність до участі в окисновально-віновних перетвореннях, у складі каталази він забезпечує відновлення Н2О2, тоді як у складі цитохрому гем виконує роль переносника електронів, змінюючи при цьому валентність заліза. У ферментах із четвертинною структурою кількість активних центрів, зазвичай, співпадає з числом субодиниць.
Активний центр простого фермента – це тривимірний утвір, здебільшого щілиноподібної форми, який утворюється сукупністю бічних радикалів амінокислотних залишків, які дуже часто знаходяться на певній відстані в лінійній послідовності поліпептидного ланцюга. Так, у молекулі лізоциму – ферменту, що забезпечує бактерицидні властивості слини, основні групи активного центра представлені амінокислотними залишками, які займають у поліпептидному ланцюзі 35, 52, 62, 63 і 101 положення. У складних ферментах активний центр може містити кофактор, а бічні радикали створюють умови для правильної конформації активного центра, орієнтації та перетворення субстрату. Проте, саме білок у складному ферменті організовує ефективне функціонування кофактора. Так, гем у комплексі з глобіном виявляє свою схильність до участі в окисновально-віновних перетвореннях, у складі каталази він забезпечує відновлення Н2О2, тоді як у складі цитохрому гем виконує роль переносника електронів, змінюючи при цьому валентність заліза. У ферментах із четвертинною структурою кількість активних центрів, зазвичай, співпадає з числом субодиниць.
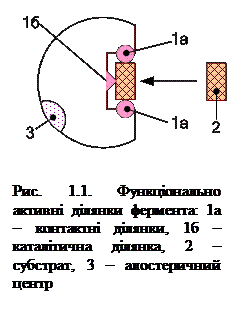
Оскільки субстрат сполучається з активним центром у кількох точках і це, своєю чергою, забезпечує високу вибірковість зв’язування (виявляється відповідність субстрату й активного центра) і орієнтацію субстрату, необхідного для каталізу, тому в активному центрі умовно виділяють якірні (або контактні) ділянки,які забезпечують вибір субстрату та його приєднання нековалентними зв’язками, а також каталітичну ділянку, яка забезпечує вибір шляху хімічного перетворення певного субстрату, тобто, бере безпосередню участь у синтезі або розриві зв'язків субстрату з утворенням продукту (рис. 1.1). Найчастіше до складу активних центрів входять такі амінокислоти як сер, гіс, глу, асп, цис-SH, тир, три, ліз,гідрофобні ланцюги аліфатичних амінокислот і ароматичне кільце фенілаланіну.
Крім активного центра, у ферментах може знаходитися алостеричний центр (або центри) (грец. allos – інший і steros – просторовий, структурний)(рис. 5.1). Він просторово розділений з активним центром і являє собою ділянку молекули фермента, з якою зв'язуються так звані модулятори або алостеричні ефектори, які за своєю природою різняться з субстратами. Вони змінюють третинну, а іноді й четвертинну структуру молекули фермента, конформацію активного центра, спричинюючи в такий спосіб пришвидшення (активатори) або сповільнення (інгібітори) ферментативної реакції. Такими ефекторами можуть бути гормони та їх похідні, метаболіти, медіатори тощо.
1.1.5. Специфічність дії ферментів.Специфічність – одна з найважливіших властивостей ферментів, яка визначає біологічну значимість цих білкових молекул і істотно відрізняє їх від небілкових каталізаторів. В основі специфічності лежить відповідність стеричної структури субстрату й активного центра фермента, внаслідок чого даний фермент з безлічі речовин, які є в клітині, приєднує лише певний субстрат.
Розрізняють субстратну та каталітичну специфічності фермента. Субстратна специфічність – це здатність фермента взаємодіяти з одним або кількома субстратами. Вона може бути абсолютною, груповою та стереоспецифічністю. Фермент з абсолютною субстратною специфічністю каталізує перетворення лише одного субстрату з певною структурою. Будь-які модифікації (зміни) у структурі субстрату роблять його недоступними для дії ферменту. Прикладом можуть слугувати аргіназа, яка каталізує реакцію розщеплення аргініну на сечовину та орнітин, та уреаза, яка каталізує гідроліз сечовини з утворенням вуглецю оксиду (ІІ) та аміаку. Сахараза гідролізує тільки сахарозу, а на інші дисахариди не діє.
Переважна більшість ферментів каталізує однотипні реакції з невеликою кількістю (групою) структурно подібних субстратів із характерним типом зв’язку, тобто вони володіють груповою субстратною специфічністю. так, панкреатична ліпаза гідролізує жири в дванадцятипалій кишці, каталізуючи перетворення будь-якої молекули жиру до моноацилгліцеролу та двох молекул вищих жирних кислот, розриваючи ефірний зв'язок біля a-атомів вуглецю гліцеролу, незалежно від того, які жирні кислоти входять до складу молекули жиру. Протеолітичні ферменти (пепсин, трипсин, хімотрипсин) теж володіють груповою субстратною специфічністю, гідролізуючи пептидні зв’язки, утворені різними амінокислотними залишками.
Ферменти зі стереохімічною специфічністю діють лише на певні стереоізомери. Наприклад, більшість моносахаридів і продуктів їх обміну в організмі людини належать до D-стереоізомерів; ферменти, які здійснюють їх метаболізм, не володіють специфічністю до L-форм. Білки людини складаються з амінокислот L-ряду, тому більшість ферментів, які забезпечують перетворення амінокислот, володіють стереоспецифічністю до L-амінокислот. Якщо сполука існує в формі цис- і трансізомерів, то тільки одна з них може служити субстрат ом для дії ферменту, наприклад, фумараза каталізує перетворення фумарої кислоти (транс-ізомер), але не діє на малеїнову кислоту (цис-ізомер). Виключення становлять лише епімерази (рацемази), які каталізують перетворення оптичних ізомерів. Стереоспецифічність до a- та b-глікозидних зв’язків можна розглянути на прикладі амілази, яка діє лише на a-глікозидні зв’язки, що дозволяє гідролізувати крохмаль і глікоген. Клітковина, хоч і теж складається з залишків глюкози, не гідролізується в організмі людини, оскільки містить у своєму складі b-глікозидні зв’язки, для гідролізу яких в організмі людини немає відповідних ферментів.
Каталітична специфічність – це такий вид специфічності, коли фермент каталізує перетворення приєднаного субстрату за одним із кількох можливих шляхів. Так, наприклад, молекула глюкозо-6-фосфату в гепатоцитах слугує субстратом для 4 різних ферментів: фосфоглюкомутази, глюкозо-6-фосфатфосфатази, фосфоглюкоізомерази та глюкозо-6-фосфатдегідрогенази. Завдяки особливостям будови каталітичних ділянок цих ферментів відбувається перетворення цього субстрату на 4 різних продукти.
1.1.6. Ізоферменти.Під множинними молекулярними формами ферментів розуміють низку форм одного і того ж фермента, які каталізують одну і ту ж реакцію, але відрізняються за місцем локалізації в організмі, складом, а отже, і за деякими властивостями: швидкістю переміщення в електричному полі, оптимумом рН, імунологічними характеристиками тощо.
Група споріднених ферментів, які мають кілька форм одного і того ж фермента з четвертинною структурною організацією і відмінних один від одного якісним і кількісним складом субодиниць, отримала назву ізоферментів. У залежності від кількості наявних поліпептидних субодиниць, а також від будови фермента (димер, тетрамер, полімер) можлива різна кількість комбінацій поліпептидних ланцюгів або різна кількість ізоферментів. Властивості ізоферментів обумовлені властивостями субодиниць, які входять до їх складу.
Зазвичай органи характеризуються різним кількісним складом того чи іншого ізофермента. Відмінності в будові поліпептидних ланцюгів та особливості поєднання цих ланцюгів у молекулі ізофермента обумовлюють відмінності в його загальному електричному заряді що, у свою чергу, забезпечує різну електрофоретичну рухливість. Це дозволяє фракціонувати ізоферменти за допомогою електрофорезу.
 У даний час ізоферменти виявлені у понад 100 ферментів. Прикладом ізоферментів може слугувати лактатдегідрогеназа (ЛДГ) – фермент, що каталізує утворення й окиснення молочної кислоти. За допомогою електрофореграми виявлено 5 його ізоформ (рис. 1.2).
У даний час ізоферменти виявлені у понад 100 ферментів. Прикладом ізоферментів може слугувати лактатдегідрогеназа (ЛДГ) – фермент, що каталізує утворення й окиснення молочної кислоти. За допомогою електрофореграми виявлено 5 його ізоформ (рис. 1.2).
Кожна ізоформа містить 4 субодиниці двох типів – «Н» (від англ. heart - серце) та «М» (від англ. muscle – м’яз) і складається з таких протомерів: ЛДГ1 = Н4; ЛДГ2 = МН3; ЛДГ3 = М2Н2; ЛДГ4 = М3Н1; ЛДГ5 = М4.
Для кожної тканини притаманне своє співвідношення ізоформ. Так, ЛДГ1 та ЛДГ2 переважають у органах, які характеризуються аеробним метаболізмом - серці, мозку, нирках, підшлунковій залозі та еритроцитах; ЛДГ3 – у легенях, селезінці, лімфатичних вузлах; ЛДГ4 та ЛДГ5 – в органах з активним анаеробним обміном – печінці та скелетних м’язах.
субодиниці ЛДГ відрізняються за спорідненістю до субстратів. Субодиниця „М” краще каталізує перетворення піровиноградної кислоти до молочної. Субодиниця „Н” є менш специфічною за своєю дією та каталізує взаємоперетворення a-кето- та a-гідроксимасляної кислоти в реакції:
СН3-СН2-СО-СООН+ НАДН2 → СН3-СН2-СНОН-СООН + НАД+
Це дозволяє відрізнити ізоферменти, які складаються, основним чином, з субодиниць „Н” (ЛДГ1, ЛДГ2) від ізофермента, побудованого з „М”-субодиниць (ЛДГ5) на підставі дослідження їх активності в реакції, яка визначається як реакція дегідрогенази a-гідроксимасляної кислоти та відбувається за участю ЛДГ1.
Вивчення ізоферментного спектра широко використовується в клініці для діагностики органічних і функціональних уражень органів і тканин, встановлення чіткої локалізації патологічного процесу. Так, розподіл ізоферментів ЛДГ у сироватці крові за умов норми має наступний вигляд: ЛДГ2 > ЛДГ1 > ЛДГ3 > ЛДГ4 > ЛДГ5. При інфаркті міокарда ЛДГ1 > ЛДГ2 > ЛДГ3 > ЛДГ4 > ЛДГ5 - помітна характерна зміна „розташування” ЛДГ1 та ЛДГ2: активність ЛДГ1 переважає над активністю ЛДГ2. При гемолітичній анемії ЛДГ2 + ЛДГ1 > ЛДГ3 + ЛДГ4 + ЛДГ5, загальна активність ЛДГ при цьому в 2 – 5 разів перевищує норму. При мегалобластних анеміях активність перевищує норму в 10 – 50 разів. Для захворювань м’язів, печінки, утворення пухлин розподіл ізоформ має наступний вигляд: ЛДГ4 = ЛДГ5 >ЛДГ2 > ЛДГ1 > ЛДГ3.
Крім ізоферментів лактатдегідрогенази велике клінічне значення мають ізоферменти креатинфосфокінази (КК). Цей фермент є димером і побудований з двох основних субодиниць – В та М, з них утворюються три комбінації цих субодиниць і, як наслідок, триізоферменти КК. У мозку переважає ізоформа ВВ, у скелетних м’язах – ММ, а у серцевому м’язі – ВМ.
Визначення активності ізоферменту МВ найчастіше використовують для підтвердження діагнозу інфаркту міокарда. Підвищену активність цієї ізоформи виявляють вже на 4 – 6 годину після відчуття болю, пов’язаного з інфарктом, тоді як загальна активність КК може значно зростати при будь-якому пошкодженні скелетних м’язів (метаболічні порушення, запалення, ішемія тощо). Зростання активності ВВ-ізоформи супроводжує пошкодження мозку (крововилив у мозок, церебральний емболізм), деякі ракові захворювання. Проте, воно не має великого діагностичного значення.
1.1.7. Рівні структурної організації ферментів. Функціональні ферментативні системи. Більшість ферментів має чотири рівні структурної організації (первинну, вторинну, третинну і четвертинну), тобто є олігомерами, які складаються з протомерів. Кожна із субодиниць або окремі їх частини відіграють певну роль у процесі функціонування ферменту. Прості (однокомпонентні) ферменти здійснюють ферментативне перетворення субстрату з участю власне білкової молекули. Безпосередню участь у реакції бере не весь поліпептидний ланцюг ферменту, а тільки незначна його частина, що близько прилягає до субстрату – активний центр. У ферментативну реакцію включається тільки декілька просторово наближених до нього залишків амінокислот. За утворення активного центру ферменту, як і за його каталітичну дію, відповідає третинна структура білкової молекули. Отже, при порушенні третинної структури (денатурація) роз’єднуються просторово розміщені амінокислотні залишки і, як наслідок, фермент втрачає активність.
Активний центр складних (двокомпонентних) ферментів, як відомо, містить у своєму складі як кофермент, так і ту частину апоферменту, що просторово прилягає до нього. Кофермент при цьому може відповідати за утворення зв’язку із субстратом, формування третинної або четвертинної структури апоферменту і каталітичне перетворення субстрату.
Кожна клітина організму має свій специфічний набір ферментів. У клітині дія кожного ферменту не індивідуальна, а тісно пов’язана з іншими ферментами. Так із окремих ферментів формуються поліферментативні (мультиферментативні) комплекси, що каналізують послідовності спряжених біохімічних реакцій.
Можна умовно виділити наступні види організації поліферментативних систем: функціональна, структурно-функціональна і змішана.
Функціональна організація характеризується тим, що окремі ферменти, об’єднанні в поліферментну систему, виконують певну функцію, з допомогою метаболітів, які дифундують від одного ферменту до іншого. Причому в функціонально організованих поліферментативних системах продукт реакції першого ферменту в ланцюгу служить субстратом для наступного тощо.
Часто метаболіт може виконувати об’єднуючу роль між різними поліферментними системами, якщо він є загальним для індивідуальних ферментів різних поліферментативних ланцюгів, кожен з яких виконує свої біохімічні функції в клітині. Такі метаболіти об’єднують ферментативні системи навіть локалізовані в різних органоїдах клітини в єдину метаболічну карту клітини.
Прикладом функціональної організації поліферментної системи служить гліколіз, тобто розпад глюкози. Всі ферменти гліколізу знаходяться в розчиненому стані. Кожна реакція каталізується окремими ферментами. Зв’язуючою ланкою тут виступають метаболіти. Положення кожного ферменту в ланцюгу встановлюється за їх спорідненістю до субстрату.
Структурно-функціональна організація полягає в тому, що ферменти утворюють структурні системи з певною функцією при допомомозі фермент-ферментативних (білок-білкових) взаємодій. Таким чином формуються поліферментативні надмолекулярні комплекси. До них відноситься, наприклад, поліферментний комплекс піруватдегідрогеназа, який складається із декількох ферментів, що беруть участь в окисненні пірувату, або синтетаза жирних кислот, яка складається із семи структурно зв’язаних ферментів. Такі поліферментативні комплекси досить міцні і важко розпадаються на окремі ферменти.
Низка таких мультиферментативних комплексів іноді називають ферментними ансамблями, які структурно пов’язані з певними органелами (рибосоми, мітохондрії) або з біомембраною і становлять високоорганізовані надмолекулярні системи (наприклад, тканинне дихання, пов’язане з перенесенням електронів від субстрату до кисню через систему дихальних ферментів).
Змішаний тип організації поліферментативних систем становить комбінацію обох типів організації, тобто частина поліферментної системи має структурну організацію, частина – функціональну (такі ферменти називають поліфункціональними). Прикладом такої організації служить поліферментна система циклу Кребса, де частина ферментів об’єднана в структурний комплекс (альфа-кетоглутаратдегідрогеназний комплекс), а частина з’єднана одна з одною функціонально за допомогою метаболітів.
Отже, розрізняють такі типи мультиферментативних систем у клітині:
1. Розчинні мультиферментативні системи, в яких відсутня постійна асоціація між ферментами, відбувається дифузія субстратів між окремими ферментами.
2. Мультиферментативні системи, в яких окремі ферменти сполучені між собою нековалентними зв’язками, утворюючи комплекси, які полегшують передавання субстратів та продуктів між окремими ферментами.
3. Мембранозв’язані мультиферментативні системи, в яких окремі ферменти асоційовані з ліпідним біщаром субклітинних органел (мітохондрій, ендоплазматичного ретикулуму тощо).
1.1.8. Внутрішньоклітинна локалізація ферментів. Органоспецифічність.Питання про локалізацію ферментів у клітині набуває особливої важливості, коли необхідно виділити фермент у чистому вигляді. Порівняно легко виявити локалізацію фермента методами цито- і гістохімії. Для цього тонкі зрізи тканин інкубують з відповідними субстратами, а після інкубації локалізацію продукту реакції досліджують за розвитком специфічної реакції при додаванні певних реактивів.
У препаративній ферментології часто користуються методом диференційного центрифугування гомогенатів тканин. Для цього спочатку руйнують клітинну структуру з допомогою відповідного дезінтегратора і утворена гомогенізована маса підлягає диференційному центрифугуванню при температурі 0 - 4о С. Центрифугування різними швидкостями гомогенатів різних тканин забезпечує седиментацію (осідання) субклітинних структур, що дозволяє згодом дослідити їх склад (табл.1.2).
Таблиця 1.2. Локалізація деяких ферментів усередині клітини
| Цитозоль | Мітохондрії | Лізосоми |
| Ферменти гліколізу Ферменти пентозофосфатного циклу Ферменти активації амінокислот Мультиферментний комплекс синтезу жирних кислот Ферменти катаболізму пуринових і піримідинових основ Пептидази Амінотрансферази Малатдегідрогеназа Ізоцитратдегідрогеназа (НАД-залежна) Глікогенфосфорилаза Глікогенсинтетаза | Піруватдегідрогеназний комплекс Цитратсинтетаза Ізоцитратдегідрогеназа (НАД-залежна) Малатдегідрогеназа та інші ферменти циклу Кребса Ацил-КоА-дегідрогеназа та інші ферменти окиснення жирних кислот Ферменти дихального ланцюга та окиснювального фосфорилування | Кисла фосфатаза β-глюкуронідаза α-глюкозидаза β-глюкозидаза Катепсини Кисла рибонуклеаза α-галактозидаза Лізоцим Гіалуронідаза Арилсульфатаза Колагеназа |
| Мікросомна фракція | Плазматична мембрана | Ядро |
| НАДН- і НАДФН- цитохром С-редуктази, цитохром Р450- і цитохром b5-оксидази, глюкозо-6-фосфатази, естерази, нуклеозиддифосфатази, глюкуронілтрансферази, фосфогліцерид- і тріацилгліцеридсинтетази, b-глюкуронідази Глюкозо-6-фосфатаза Рибосомні ферменти синтезу білка Ферменти, які беруть участь у реакціях гідроксилування Ферменти синтезу фосфоліпідів, тригліцеридів, а також деякі ферменти синтезу холестерину | Аденілатциклаза Лужна фосфатаза Nа+-К+-залежна АТФаза | Ферменти, які беруть участь у процесі реплікації ДНК РНК-полімераза НАД-синтетаза |
| Ендоплазматичний ретикулум | Комплекс Гольджі | |
| Глюкозо-6-фосфатаза | Галактозилтрансфераза |
Наявність визначених ферментів у внутрішньоклітинних ділянках (компартментах) обумовлює функціонування в них метаболічних шляхів – перетворення одних метаболітів на інші.
Відповідно, ферментний склад різних клітин різний. Ферменти, які виконують функцію життєзабезпечення, знаходяться в усіх клітинах організму. У процесі диференціювання клітин відбувається зміна їх ферментного складу. Так, фермент аргіназа який бере участь в утворенні сечовини, знаходиться лише в клітинах печінки, а кисла фосфатаза, яка бере участь у гідролізі моно ефірів ортофосфорної кислоти – у клітинах передміхурової залози. Це так звана органоспецифічність ферментів. Якщо говорити про вузькоспеціалізовані клітини, то ферментів, які забезпечують їх функціонування знаходиться в цих клітинах більше порівняно з іншими. Наприклад, у клітинах міокарда підвищена кількість креатинкінази та аспартатамінотрансферази, у гепатоцитах – аланінамінотрансферази, в остеобластах – лужної фосфатази тощо.
1.1.9. Одиниці активності ферментів. Активність ферментів виражають в міжнародних одиницях активності. Кількість ферменту, яка каталізує перетворення 1 мікромоля субстрату на продукт реакції за 1хв у стандартних (оптимальних) умовах з розрахунку на 1 г тканини називається міжнародною одиницею (МО). Для оцінки кількості молекул фермента серед інших білків досліджуваної тканини визначають питому активність – це кількість одиниць фермента в зразку тканини, розділену на масу білка (мг) у цій тканині. За цією величиною аналізують ступінь очистки фермента: чим менше сторонніх білків, тим вища питома активність.
У 1973 році Міжнародна біохімічна спілка запропонувала використати як одиницю активності катал (кат). Активність в 1 кат – це така кількість каталізатора, яка перетворює 1 моль субстрату на продукт за 1 секунду. Отже, 1 кат = 60×106 міжнародних одиниць. Рекомендовано використати також одиницю значно меншого масштабу - нанокатал (нкат), яка дорівнює 109 кат.
Кількість молекул субстрату, які перетворюються однією молекулою ферменту на продукт у процесі реакції за одиницю часу при повному насиченні ферменту субстратом, прийнято називати числом обертів ферменту, або молярною активністю. Ця активність виражається в каталах на 1 моль ферменту.
1.1.10. Принципи та методи виявлення ферментів у біооб’єктах.Здебільшого кількість фермента не можна виміряти в одиницях маси, тобто в міліграмах або в молях, про неї опосередковано свідчить його активність, тобто дія фермента. Іншими словами, присутність і кількість фермента виявляють за специфічністю та швидкістю реакції, яку він каталізує. Активність фермента можна визначити опосередковано: за визначенням кількості продукту, який утворився під дією ферменту, або за кількістю витраченого субстрату за одиницю часу при оптимальних умовах ферментативної реакції. Як приклад розглянемо принцип методу визначення активності a-амілази, яка каталізує гідролітичне розщеплення крохмалю з утворенням у кінцевому результаті мальтози. Активність амілази розраховують на основі кількості негідролізованого крохмалю або за кількістю утвореної мальтози.
Найчастіше активність ферментів визначають у сироватці, плазміабоклітинах крові та в сечі колориметричними, спектрофотометричними, флюориметричними, манометричними, кондуктометричними, віскозиметричними методами. У клініко-діагностичних лабораторіях для визначення активності ферментів переважно застосовують колориметричні та спектрофотометричні методи.
В основіколориметричних методів лежить вимірювання за допомогою фотоелектроколориметра інтенсивності забарвлення речовини, яка утворюється під час взаємодії субстрату або продукту ферментативної реакції зі специфічними реактивами, які додаються у пробу після припинення реакції. Наприклад, продуктом дії ЛДГ в умовах прямої реакції є піруват, який з 2,4-динітро-фенілгідразином утворює у лужному середовищі забарвлений гідразон. Інтенсивність забарвлення пропорційна кількості пірувату, а значить, і активності фермента.
Спектрофотометричні методи реєструють поглинання світла в певних ділянках спектра коферментами, субстратами або продуктами реакції. Ці методи широко застосовують для визначення активності оксидоредуктаз, зокрема НАД- та НАДФ-залежних дегідрогеназ. Перехід НАД+ або НАДФ+ з окисненої форми у відновлену супроводжується зміною поглинання в ультрафіолетовій ділянці спектра. В окисненій формі кофермент дає одну вузьку смугу поглинання при λ=260 нм, яка обумовлена наявністю аденіну в його молекулі.
 У разі відновлення кофермента поглинання світла в цій зоні дещо знижується, але з’являється інша широка смуга поглинання при λ=340 нм. Поява цієї смуги зумовлена зникненням одного подвійного зв’язку у піридиновому кільці аденіну під час його відновлення. Визначення активності ферментів за різницею спектрів поглинання окисненої та відновленої форм НАД+ (або НАДФ+) та НАДН (або НАДФН) називають тестом Варбурга (рис. 1.3).
У разі відновлення кофермента поглинання світла в цій зоні дещо знижується, але з’являється інша широка смуга поглинання при λ=340 нм. Поява цієї смуги зумовлена зникненням одного подвійного зв’язку у піридиновому кільці аденіну під час його відновлення. Визначення активності ферментів за різницею спектрів поглинання окисненої та відновленої форм НАД+ (або НАДФ+) та НАДН (або НАДФН) називають тестом Варбурга (рис. 1.3).
Читайте також:
- III. Географічна структура світового ринку позичкового капіталу
- VІ. План та організаційна структура заняття
- А) Структура економічних відносин.
- А) Товар і його властивості.
- Адміністративно – територіальний устрій і соціальна структура Слобожанщини у половині XVII – кінці XVIII століття
- Аеродинамічні властивості колісної машини
- Акти з охорони праці, що діють в організації, їх склад і структура.
- Алфавіт мови і структура програми
- Аналізатори людини та їхні властивості.
- Аналізатори людини та їхні властивості.
- АРХІВНІ ДОВІДНИКИ В СИСТЕМІ НДА: ФУНКЦІЇ ТА СТРУКТУРА
- АРХІВНІ ДОВІДНИКИ В СИСТЕМІ НДА: ФУНКЦІЇ ТА СТРУКТУРА
| <== попередня сторінка | | | наступна сторінка ==> |
| РОЗДІЛ 1. ФЕРМЕНТИ | | | Номенклатура та класифікація ферментів |
|
Не знайшли потрібну інформацію? Скористайтесь пошуком google: |
© studopedia.com.ua При використанні або копіюванні матеріалів пряме посилання на сайт обов'язкове. |