РЕЗОЛЮЦІЯ: Громадського обговорення навчальної програми статевого виховання
ЧОМУ ФОНД ОЛЕНИ ПІНЧУК І МОЗ УКРАЇНИ ПРОПАГУЮТЬ "СЕКСУАЛЬНІ УРОКИ"
ЕКЗИСТЕНЦІЙНО-ПСИХОЛОГІЧНІ ОСНОВИ ПОРУШЕННЯ СТАТЕВОЇ ІДЕНТИЧНОСТІ ПІДЛІТКІВ
Батьківський, громадянський рух в Україні закликає МОН зупинити тотальну сексуалізацію дітей і підлітків
Відкрите звернення Міністру освіти й науки України - Гриневич Лілії Михайлівні
Представництво українського жіноцтва в ООН: низький рівень культури спілкування в соціальних мережах
Гендерна антидискримінаційна експертиза може зробити нас моральними рабами
ЛІВИЙ МАРКСИЗМ У НОВИХ ПІДРУЧНИКАХ ДЛЯ ШКОЛЯРІВ
ВІДКРИТА ЗАЯВА на підтримку позиції Ганни Турчинової та права кожної людини на свободу думки, світогляду та вираження поглядів
- Гідрологія і Гідрометрія
- Господарське право
- Економіка будівництва
- Економіка природокористування
- Економічна теорія
- Земельне право
- Історія України
- Кримінально виконавче право
- Медична радіологія
- Методи аналізу
- Міжнародне приватне право
- Міжнародний маркетинг
- Основи екології
- Предмет Політологія
- Соціальне страхування
- Технічні засоби організації дорожнього руху
- Товарознавство продовольчих товарів
Тлумачний словник
Авто
Автоматизація
Архітектура
Астрономія
Аудит
Біологія
Будівництво
Бухгалтерія
Винахідництво
Виробництво
Військова справа
Генетика
Географія
Геологія
Господарство
Держава
Дім
Екологія
Економетрика
Економіка
Електроніка
Журналістика та ЗМІ
Зв'язок
Іноземні мови
Інформатика
Історія
Комп'ютери
Креслення
Кулінарія
Культура
Лексикологія
Література
Логіка
Маркетинг
Математика
Машинобудування
Медицина
Менеджмент
Метали і Зварювання
Механіка
Мистецтво
Музика
Населення
Освіта
Охорона безпеки життя
Охорона Праці
Педагогіка
Політика
Право
Програмування
Промисловість
Психологія
Радіо
Регилия
Соціологія
Спорт
Стандартизація
Технології
Торгівля
Туризм
Фізика
Фізіологія
Філософія
Фінанси
Хімія
Юриспунденкция
Вплив ліганда білку на розпад мРНК
Проте, схоже, в регуляції розпаду мРНК іноді бувають задіяні і складніші зв'язки. Прикладом є мРНК трансферину - білку, який потрібний для транспорту іонів заліза в плазмі крові і перенесення їх в клітини. (У самих же клітинах іони Fе2+ зв'язуються з іншим білком - феритином). Утворюється трансферин в гепатоцитах.
Так от, при нестачі іонів заліза в клітинах печінки стабільність трансферинової мРНК висока. Тому трансферин інтенсивно утворюється і забезпечує зв'язування заліза (у місцях руйнування еритроцитів, з продуктів харчування і т. д.). Це кінець кінцем сприяє підвищенню концентрації іонів Fе2+ і в клітинах (як у гепатоцитах, так і в інших клітинах).
При надлишку ж цих іонів в гепатоцитах стабільність трансферинової мРНК значно знижується. Відповідно, слабшає і синтез трансферину.
Як бачимо, в даному випадку на інтенсивність розпад мРНК чинить вплив не сам кодований нею білок, а ліганд, що зв'язується з цим білком.
Можливий механізм такого впливу показаний мал.
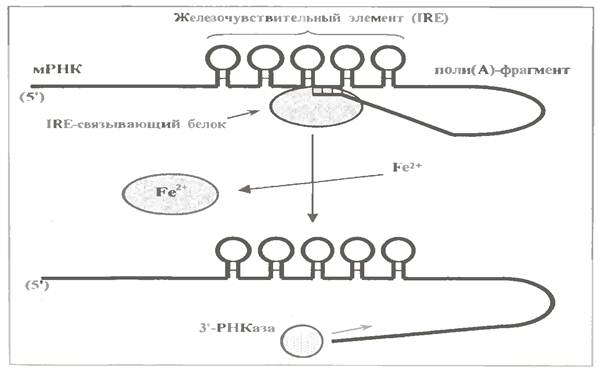
У 3´-нетрансльованій області мРНК трансферина є 5 "шпильок", що утворюють т. з. залізочутливий елемент (IRE - iron - responsive element). У відсутність іонів Fe2+ тут зв'язується спеціальний білок - IRE-зв'язуючий білок. І це якимось чином захищає пів(А) -фрагмент мРНК від негайної дії 3'-РНКази. Не виключено, що цей білок, взаємодіючи одночасно з IRE і 3´-кінцем пів(А) -фрагмента, полегшує утворення пів(А) -фрагментом петлі із захищеним 3'-концом.
Ионы же Fe2+, накопичуючись в гепатоцитах, викликають дисоціацію IRE -зв'язуючого білку. Тому пів(А) -фрагмент мРНК трансферину стає доступним для 3'-РНКази.
Читайте також:
- I визначення впливу окремих факторів
- Аденогіпофіз, його гормони, механізм впливу
- Аденогіпофіз, його гормони, механізм впливу, прояви гіпер- та гіпофункцій.
- Адміністративні методи - це сукупність прийомів, впливів, заснованих на використанні об'єктивних організаційних відносин між людьми та загальноорганізаційних принципів управління.
- Активний вплив на проблему
- Активність. Закон радіоактивного розпаду
- Аналіз відхилень від нормативів та їх впливу на прибуток
- Аналіз впливу постачальників
- Аналіз впливу факторів на зміну сумми гуртової реалізації
- Аналіз факторів впливу на обсяги виробництва суспільного продукту.
- Аналіз факторів, що впливають на цінову політику.
- Анатомо-фізіологічна перебудова організму підлітка та її вплив на його психологічні особливості й поведінку.
| <== попередня сторінка | | | наступна сторінка ==> |
| Продукти транскрипції | | |
|
Не знайшли потрібну інформацію? Скористайтесь пошуком google: |
© studopedia.com.ua При використанні або копіюванні матеріалів пряме посилання на сайт обов'язкове. |