РЕЗОЛЮЦІЯ: Громадського обговорення навчальної програми статевого виховання
ЧОМУ ФОНД ОЛЕНИ ПІНЧУК І МОЗ УКРАЇНИ ПРОПАГУЮТЬ "СЕКСУАЛЬНІ УРОКИ"
ЕКЗИСТЕНЦІЙНО-ПСИХОЛОГІЧНІ ОСНОВИ ПОРУШЕННЯ СТАТЕВОЇ ІДЕНТИЧНОСТІ ПІДЛІТКІВ
Батьківський, громадянський рух в Україні закликає МОН зупинити тотальну сексуалізацію дітей і підлітків
Відкрите звернення Міністру освіти й науки України - Гриневич Лілії Михайлівні
Представництво українського жіноцтва в ООН: низький рівень культури спілкування в соціальних мережах
Гендерна антидискримінаційна експертиза може зробити нас моральними рабами
ЛІВИЙ МАРКСИЗМ У НОВИХ ПІДРУЧНИКАХ ДЛЯ ШКОЛЯРІВ
ВІДКРИТА ЗАЯВА на підтримку позиції Ганни Турчинової та права кожної людини на свободу думки, світогляду та вираження поглядів
- Гідрологія і Гідрометрія
- Господарське право
- Економіка будівництва
- Економіка природокористування
- Економічна теорія
- Земельне право
- Історія України
- Кримінально виконавче право
- Медична радіологія
- Методи аналізу
- Міжнародне приватне право
- Міжнародний маркетинг
- Основи екології
- Предмет Політологія
- Соціальне страхування
- Технічні засоби організації дорожнього руху
- Товарознавство продовольчих товарів
Тлумачний словник
Авто
Автоматизація
Архітектура
Астрономія
Аудит
Біологія
Будівництво
Бухгалтерія
Винахідництво
Виробництво
Військова справа
Генетика
Географія
Геологія
Господарство
Держава
Дім
Екологія
Економетрика
Економіка
Електроніка
Журналістика та ЗМІ
Зв'язок
Іноземні мови
Інформатика
Історія
Комп'ютери
Креслення
Кулінарія
Культура
Лексикологія
Література
Логіка
Маркетинг
Математика
Машинобудування
Медицина
Менеджмент
Метали і Зварювання
Механіка
Мистецтво
Музика
Населення
Освіта
Охорона безпеки життя
Охорона Праці
Педагогіка
Політика
Право
Програмування
Промисловість
Психологія
Радіо
Регилия
Соціологія
Спорт
Стандартизація
Технології
Торгівля
Туризм
Фізика
Фізіологія
Філософія
Фінанси
Хімія
Юриспунденкция
ВНУТРІШНЬОКЛІТИННІ СТРУКТУРИ.
Рибосоми.Рибосоми є місцем синтезу білка. На електронних мікрофотографіях вони мають вигляд частинок, що лежать у цитоплазмі. Рибосоми бактерій мають розміри 16 х 18 нм. Приблизно 80-85 % всієї бактеріальної РНК міститься в рибосомах. Складаються з білка (35-40 %) та РНК (60-65 %). Оскільки інтактні (цілі, незруйновані) рибосоми бактерій при ультрацентрифугуванні осідають із швидкістю близько 70 одиниць Сведберга (S), їх називають 70 S -рибосомами. Цитоплазматичні рибосоми еукаріот більші — 80S. З мітохондрій еукаріот (наприклад, з мітохондрій дріжджів) можна також виділити рибосоми, які за розмірами схожі на бактеріальні.
Бактеріальна 70 S -рибосома складається з двох субодиниць — 30S- та 50 S -субодиниці (рис. 2.8). Бактеріальна клітина містить від 5 000 до 50 000 рибосом. їх кількість тим більша, чим швидше росте клітина. Під час активного синтезу білка на електронних мікрофотографіях можна бачити правильні ланцюжки рибосом. Це рибосоми, нанизані, як намисто, на ланцюги матричної РНК; їх називають полірибосомами або полісомами.
Різниця між рибосомами бактерій (70S) та еукаріот (80S) має вирішальне значення для боротьби з інфекційними хворобами. Деякі антибіотики частково чи повністю пригнічують синтез білка, який відбувається на 708-рибосомах, і не порушують функцій 80S-рібосом.
Вакуолі. Газові вакуолі (аеросоми) характерні для водних бактерій, особливо пурпурових і зелених сіркобактерій, мешканців мулів, окремих ґрунтових бактерій. Газові вакуолі складаються з газових бульбашок, розміщених паралельними рядами, утворюючи стільникоподібну структуру. Газові бульбашки — це порожнисті циліндри, оточені одношаровою білковою мембраною. У цій білковій мембрані гідрофобні амінокислоти повернуті всередину циліндра, а гідрофільні — назовні. Таке розміщення амінокислот перешкоджає проникненню води в бульбашку. Циліндри заповнені газом, склад якого є аналогічним складу газу навколишнього середовища. Аеросоми є регуляторами плавучості бактерій. Вони дають можливість бактеріям, які не мають джгутиків, здійснювати вертикальний рух у водоймах і капілярах ґрунту і таким чином займати найвигідніше положення відносно джерела світла, концентрації розчиненого кисню і поживних речовин, тобто газові вакуолі у бактерій мають пристосувальне значення.
Щодо еукаріотних клітин, то вони містять велику вакуоль, оточену одношаровою мембраною. її функції точно не встановлені. Показано, що в ній містяться гідролітичні ферменти, поліфосфати, ліпіди, низькомолекулярні клітинні інтермедіати та іони металів. Ймовірно, що вакуоль є резервуаром для зберігання поживних речовин і гідролітичних ферментів.
Карбоксисоми.Ці структури виявлені в клітинах ціанобактерій, деяких пурпурових бактерій, нітрифікуючих бактерій. Вони являють собою чотири- чи шестигранні включення діаметром до 500 нм, оточені одношаровою білковою мембраною, містять ферменти фіксації вуглекислого газу в циклі Кальвіна.
Магнітосоми.Виявлені в клітинах бактерій, яким притаманний магнітотаксис. Це частинки Fe3O4, оточені мембраною, різні за формою, кількістю і характером розміщення в клітині.

Рис.2.8. Схема будови бактеріальної рибосоми
Запасні речовини.У багатьох мікроорганізмів у певних умовах середовища відкладаються речовини, які можна розглядати як запасні — полісахариди, жири, лоліфосфати та сірка. Ці речовини накопичуються, якщо в поживному середовищі містяться певні вихідні сполуки, але разом з тим ріст мікроорганізмів обмежений чи взагалі неможливий із-за браку окремих компонентів живлення чи присутності інгібіторів. Запасні речовини містяться в клітині в осмотично інертній формі — вони нерозчинні у воді. В умовах, сприятливих для росту, коли в цих запасних речовинах виникає потреба, вони знову залучаються до метаболізму. Запасні полісахариди, нейтральні жири, полі-в-гідроксимасля-на кислота можуть бути джерелами вуглецю і енергії, тому за відсутності зовнішніх джерел енергії вони можуть продовжити термін існування клітини, а у спороутворювальних видів створити умови для утворення спор навіть за відсутності екзогенних субстратів. Поліфосфати можуть бути резервним джерелом фосфору, а сірка — потенційним донором електронів.
У деяких мікроорганізмів за допомогою кольорової реакції з розчином Люголю вдається ідентифікувати крохмаль (синє забарвлення) або глікоген (коричневе забарвлення). Запасні полісахариди, на відміну від полісахаридів клітинної стінки, утворюються з а-глюкози, молекули глюкози з єднані 1,4-а-зв'яз-ками. Завдяки а-зв'язкам поліглюкозні ланцюги не витягнуті в довжину, а закручені гвинтоподібно. Крохмалеподібна сполука гранульоза є специфічною запасною речовиною у бактерій роду Clostridium. Глікоген (тваринний крохмаль) схожий на амілопектин, але його ланцюги ще більш розгалужені (за рахунок утворення 1,6-зв'язків). У бактерій глікоген зустрічається частіше, ніж крохмаль. Він також виявлений у дріжджів і грибів.
У клітинах мікроорганізмів часто зустрічаються гранули та краплини жиру. їх можна фарбувати ліпофільними барвниками — Суданом III та Суданом чорним, при цьому вони стають видимими через мікроскоп. Без забарвлення їх можна побачити через мікроскоп завдяки тому, що вони сильно заломлюють світло.
Гранули багатьох бактерій складаються з полі-в-гідроксимасляної кислоти. Це поліефір, що містить близько 60 залишків в-гідроксибутирату. Частка цієї речовини в сухій біомасі може досягати 80 % . Полі-в-гідроксимасляну кислоту утворюють аеробні бактерії, а також ціанобактерії та анаеробні фототрофні бактерії. Утворення полі-в-гідроксимасляної кислоти спостерігається у аеробів в умовах браку кисню.
Нейтральні жири (тригліцериди) в особливо великих кількостях відкладаються у вакуолях дріжджів і грибів. Воски (складні ефіри жирних кислот і спиртів з довгим ланцюгом) виявлені у мікобактерій (можуть містити до 40 % восків), нокардій та актиноміцетів. Вміст запасних жирів визначається складом поживного середовища (високим співвідношенням C/N), і ці жири можуть бути виділені безпосередньо з клітин.
Гриби, дріжджі, багато бактерій і зелені водорості здатні накопичувати фосфорну кислоту у вигляді гранул поліфосфату. Такі гранули вперше були описані у бактерій Spirillum volutans. тому їх часто називають волютиновими гранулами (волютином).
У багатьох бактерій, які окиснюють сульфід до сульфату, сірка тимчасово зберігається у вигляді кульок, що сильно заломлюють світло. Кількість сірки, яка може накопичитись, залежить від вмісту сірководню в навколишньому середовищі: за відсутності сірководню сірка окиснюється до сульфату. Для аеробних сіркових бактерій сірка є джерелом енергії, а для анаеробних фототрофних пурпурових бактерій — донором електронів. Нуклеоїд. Питання про наявність ядра у бактерій було дискусійним упродовж багатьох десятиліть. Малі розміри клітин, недосконалість методів дослідження утруднювало виявлення ядерних структур у бактерій, хоча не було сумнівів щодо наявності у них спадкового апарата, оскільки при розмноженні клітини одного виду бактерій давали нащадків того самого виду. Сучасні електронно-мікроскопічні та генетичні дослідження дали змогу встановити, що бактерії мають структури, аналогічні ядрам клітин еукаріот, але відрізняються від них рядом особливостей:
1) ядра бактерій не мають ядерної оболонки (мембрани), і ДНК перебуває в безпосередньому контакті з цитоплазмою;
2) відсутній поділ на хромосоми, і нитка ДНК являє собою аналог хромосоми еукаріот і називається бактеріальною хромосомою(в клітині може бути кілька її копій);
3) відсутній мейоз і мітоз.
У зв'язку з цим ядерний апарат бактерій називають бактеріальним ядром, або нуклеоїдом. Встановлено, що бактеріальна хромосома має форму замкнутого кільця. Це гігантська молекула ДНК з молекулярною масою 109 дальтон. Вона має різну довжину у різних бактерій: у мікоплазм — 0,25, у ціанобактерій — до 3 мм. У клітинах прокаріот може бути кілька нуклеоїдів і кілька копій хромосоми (у Bacillus subtllis — від 2 до 9 хромосом у кількох нуклеоїдах, yAzotobacter vinelandii — близько 40 хромосом в одному нуклеоїді).Хромосоми бактерій — це високоупорядковані структури двох типів: зв'язані з елементами оболонки та вільні з коефіцієнтами седиментації відповідно 3200-7000S та 1600-2000S. ДНК у цих структурах перебуває в суперспіралізованому стані і утворює 20-140 петель, з'єднаних з щільною центральною ділянкою, яка складається з РНК і є відповідальною за підтримання компактної форми (рис. 2.9.).
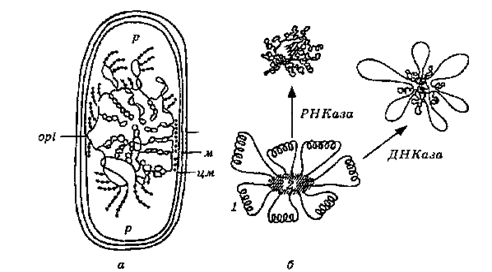
Рис. 2.9. Модель організації нуклеоїду (а) та будова компактної хромосоми Escherichia coli (б):
1 — петля хромосоми; 2 — РНК,що з'єднує петлі; орі — точка origin на хромосомі бактерії; р — рибосоми; зм — зовнішня мембрана; м — муреїн; цм — цитоплазматична мембрана
Хромосома бактерій завжди зв'язана з мембраною (кількість місць зв'язку може досягати 20 і більше) безпосередньо із специфічними мембранними білками або через рибосоми, зв'язані з мембраною.
На відміну від еукаріот, у прокаріот не спостерігається суттєвих змін у стані нуклеоїду в процесі клітинного циклу, за винятком його ущільнення перед спороутворенням. У бактеріальній клітині синтез ДНК відбувається безперервно і приблизно 1-3 % сухої маси клітини припадає на ДНК.
Читайте також:
- Господарський комплекс національної економіки: основи формування та особливості структури.
- Заголовок – назва словесного твору, яка відображає його основну ідею і є важливим елементом його структури.
- Закономірності, які розкривають характеристику функціонування і розвитку управлінської підструктури.
- Залежно від підрозділу, діяльність якого покладена в основу виробничої структури, розрізняють цехову, безцехову, корпусну та комбінатську виробничі структури.
- ЗМІСТОВИЙ МОДУЛЬ І. Внутрішньоклітинні паразити – рикетсії та хламідії
- Мережева модель організовує дані у вигляді мережевої структури. Структура називається мережевою, якщо у відносинах між даними породжений елемент має більш одного початкового.
- Мистецтво як феномен культури. Генезис мистецтва, формування його видової структури. Поняття про актуальний вид мистецтва
- Міжнародні та регіональні економічні структури.
- Основні характеристики і показники виробничої структури.
- Поняття про системи та структури. Типи систем. Типи зв’язків між елементами систем
- Поняття та елементи ринкової інфраструктури.
| <== попередня сторінка | | | наступна сторінка ==> |
| Мембранні утворення у прокаріот. | | | Характеристика спороутворювальних бактерій. |
|
Не знайшли потрібну інформацію? Скористайтесь пошуком google: |
© studopedia.com.ua При використанні або копіюванні матеріалів пряме посилання на сайт обов'язкове. |