РЕЗОЛЮЦІЯ: Громадського обговорення навчальної програми статевого виховання
ЧОМУ ФОНД ОЛЕНИ ПІНЧУК І МОЗ УКРАЇНИ ПРОПАГУЮТЬ "СЕКСУАЛЬНІ УРОКИ"
ЕКЗИСТЕНЦІЙНО-ПСИХОЛОГІЧНІ ОСНОВИ ПОРУШЕННЯ СТАТЕВОЇ ІДЕНТИЧНОСТІ ПІДЛІТКІВ
Батьківський, громадянський рух в Україні закликає МОН зупинити тотальну сексуалізацію дітей і підлітків
Відкрите звернення Міністру освіти й науки України - Гриневич Лілії Михайлівні
Представництво українського жіноцтва в ООН: низький рівень культури спілкування в соціальних мережах
Гендерна антидискримінаційна експертиза може зробити нас моральними рабами
ЛІВИЙ МАРКСИЗМ У НОВИХ ПІДРУЧНИКАХ ДЛЯ ШКОЛЯРІВ
ВІДКРИТА ЗАЯВА на підтримку позиції Ганни Турчинової та права кожної людини на свободу думки, світогляду та вираження поглядів
- Гідрологія і Гідрометрія
- Господарське право
- Економіка будівництва
- Економіка природокористування
- Економічна теорія
- Земельне право
- Історія України
- Кримінально виконавче право
- Медична радіологія
- Методи аналізу
- Міжнародне приватне право
- Міжнародний маркетинг
- Основи екології
- Предмет Політологія
- Соціальне страхування
- Технічні засоби організації дорожнього руху
- Товарознавство продовольчих товарів
Тлумачний словник
Авто
Автоматизація
Архітектура
Астрономія
Аудит
Біологія
Будівництво
Бухгалтерія
Винахідництво
Виробництво
Військова справа
Генетика
Географія
Геологія
Господарство
Держава
Дім
Екологія
Економетрика
Економіка
Електроніка
Журналістика та ЗМІ
Зв'язок
Іноземні мови
Інформатика
Історія
Комп'ютери
Креслення
Кулінарія
Культура
Лексикологія
Література
Логіка
Маркетинг
Математика
Машинобудування
Медицина
Менеджмент
Метали і Зварювання
Механіка
Мистецтво
Музика
Населення
Освіта
Охорона безпеки життя
Охорона Праці
Педагогіка
Політика
Право
Програмування
Промисловість
Психологія
Радіо
Регилия
Соціологія
Спорт
Стандартизація
Технології
Торгівля
Туризм
Фізика
Фізіологія
Філософія
Фінанси
Хімія
Юриспунденкция
Сумація збудження і гальмування нейронами ЦНС.
Оскільки, поріг деполяризації мембрани аксонного горбика складає 10-15 мВ, а амплітуда одиночного ЗПСП дорівнює 1 мВ, для виникнення ПД на мембрані аксонного горбика необхідне додавання (сумація) 10-15 ЗПСП: лише тоді деполяризація мембрани дійде до критичного рівня (Екр). В залежності від умов винекнення розрізняють розрізняють два вида сумацій: просторову та часову. Механізм їх розвитку наступний:
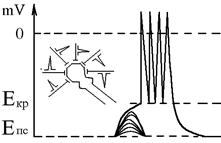 1. Просторова сумація – виникає, якщо на тілі одного нейрона одночасно функціонує декілька збудливих синапсів: в кожному із них виникають ЗПСП, які за допомогою місцевих струмів поширюються на мембрану аксонного горбика і там відбувається їх додавання (сумація). Якщо в результаті сумації ЗПСП на мембрані аксонного горбика її деполяризація доходить до критичного рівня, виникає серія ПД – ритмічний розряд нейрона (РРН). Частота ПД тим більша, чим швидше деполяризація дійде до критичного рівня, тобто від інтенсивності сумації ЗПСП.
1. Просторова сумація – виникає, якщо на тілі одного нейрона одночасно функціонує декілька збудливих синапсів: в кожному із них виникають ЗПСП, які за допомогою місцевих струмів поширюються на мембрану аксонного горбика і там відбувається їх додавання (сумація). Якщо в результаті сумації ЗПСП на мембрані аксонного горбика її деполяризація доходить до критичного рівня, виникає серія ПД – ритмічний розряд нейрона (РРН). Частота ПД тим більша, чим швидше деполяризація дійде до критичного рівня, тобто від інтенсивності сумації ЗПСП.
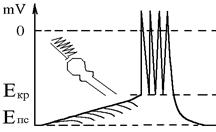 2. Часова сумація – відбувається на постсинаптичній мембрані кожного збудливого синапса, якщо частота, з якою поширюються ПД по постсинаптичній мембрані (та виникають ЗПСП на постсинаптичній) така, що кожен наступний ЗПСП починається тоді, коли ще не скінчився попередній (частота ПД має бути не менше 66 Гц, так як тривалість ЗПСП 15 мс). Відповідна сумація ЗПСП відбувається й на мембрані аксонного горбика Ù якщо вона доходить до Екр Ù серія ПД. Частоту ПД тут також визначає інтенсивність сумації.
2. Часова сумація – відбувається на постсинаптичній мембрані кожного збудливого синапса, якщо частота, з якою поширюються ПД по постсинаптичній мембрані (та виникають ЗПСП на постсинаптичній) така, що кожен наступний ЗПСП починається тоді, коли ще не скінчився попередній (частота ПД має бути не менше 66 Гц, так як тривалість ЗПСП 15 мс). Відповідна сумація ЗПСП відбувається й на мембрані аксонного горбика Ù якщо вона доходить до Екр Ù серія ПД. Частоту ПД тут також визначає інтенсивність сумації.
До сумацій здатні не тільки ЗПСП, а й ГПСП Ù ГПСП також додається на тілі нейрона за допомогою часової та просторової сумації. В один і той самий час на тілі нейрона функціонують тисячі збуджуючих та гальмівних синапсів. Взаємодія збудження та гальмування на тілі кожного окремого нейрона відбувається шляхом сумації (просторової та часової). В залежності від переважання сумації ЗПСП чи ГПСП нейрон може перебувати в трьох станах:
- збудження – характеризується генерацією ПД на мембрані аксонного горбика Ù в результаті переважання сумації ЗПСП, деполяризація мембрани дійшла до критичного рівня: чим інтенсивніше протікає сумація ЗПСП, тим швидше деполяризація доходить до Екр, тим частіше ПД в РРН (тобто, тим сильніше збудження нейрона).
- полегшення – характеризується переважанням сумації ЗПСП над ГПСП, але деполяризація мембрани аксонного горбика не доходить до Екр., ΔЕ зменшується, тобто, збудливість нейрона підвищується і наступає стан полегшення.
- гальмування - характеризується переважанням сумації ГПСП над ЗПСП, внаслідок чого величина мембранного потенціалу аксонного горбика підвищується (гіперполяризація мембрани) Ù збільшення порогу деполяризації Ù зменшення збудливості нейрона (гальмування).
Таким чином, за допомогою сумації збудження та гальмування (ЗПСП та ГПСП), кожен нейрон ЦНС здійснює обробку інформації – аналіз аферентних сигналів і синтез РРН (еферентного сигнала). В цьому і полягає фізіологічна роль процесів сумації.
Види нейронних ланцюгів ЦНС, їх роль посиленні і подовженні біологічно важливих сигналів.
Збудження і гальмування в ЦНС можуть поширюватись по таким нейронним ланцюгам:
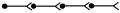 1. Лінійні – кожен попередній нейрон передає інформацію наступному. При цьому передача інформації здійснюється:
1. Лінійні – кожен попередній нейрон передає інформацію наступному. При цьому передача інформації здійснюється:
а) по аксонах нейронів (швидко, точно, без змін);
б) через центральні синапси (тут проходить не тільки передача, а й переробка інформації шляхом сумації ЗПСП і ГПСП).
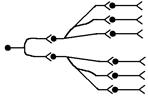 2. Дивергентні (ті що розходяться) – кожен попередній нейрон передає інформацію багатьом наступним внаслідок дивергенції (розгалуження) аксона. За участю ланцюгів такого роду в ЦНС проходить іррадіація (широке поширення) збудження, що має значення для посилення біологічно важливих сигналів.
2. Дивергентні (ті що розходяться) – кожен попередній нейрон передає інформацію багатьом наступним внаслідок дивергенції (розгалуження) аксона. За участю ланцюгів такого роду в ЦНС проходить іррадіація (широке поширення) збудження, що має значення для посилення біологічно важливих сигналів.
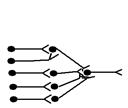 3. Конвергентні (ті що сходяться) – такі ланцюги утворюються, коли один нейрон отримує інформацію від багатьох попередніх. За участю таких ланцюгів здійснюються процеси просторової сумації на тілі нейрона.
3. Конвергентні (ті що сходяться) – такі ланцюги утворюються, коли один нейрон отримує інформацію від багатьох попередніх. За участю таких ланцюгів здійснюються процеси просторової сумації на тілі нейрона.
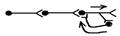 4. Кільцеві – утворюються, якщо аксон нейрона дає коллатераль, яка передає інформацію вставному нейрону, який, в свою чергу, утворює кільце, передаючи інформацію “першому” нейрону. Якщо вставний нейрон є гальмівним, то розвивається зворотнє гальмування (наприклад, мотонейронів за участю клітин Реншоу); якщо ж вставний нейрон є збуджуючим, то такі ланцюги забезпечують розвиток рефлекторної післядії – за рахунок рециркуляції імпульсів збудження по кільцевому ланцюзі, адекватна рефлекторна реакція продовжується протягом деякого часу після закінчення дії подразника. Таким чином проходить подовження біологічно важливих сигналів в ЦНС.
4. Кільцеві – утворюються, якщо аксон нейрона дає коллатераль, яка передає інформацію вставному нейрону, який, в свою чергу, утворює кільце, передаючи інформацію “першому” нейрону. Якщо вставний нейрон є гальмівним, то розвивається зворотнє гальмування (наприклад, мотонейронів за участю клітин Реншоу); якщо ж вставний нейрон є збуджуючим, то такі ланцюги забезпечують розвиток рефлекторної післядії – за рахунок рециркуляції імпульсів збудження по кільцевому ланцюзі, адекватна рефлекторна реакція продовжується протягом деякого часу після закінчення дії подразника. Таким чином проходить подовження біологічно важливих сигналів в ЦНС.
Загальну схему посилення та подовження біологічно важливих сигналів в ЦНС можна подати в наступному вигляді:
Біологічно важливий аферентний сигнал в ЦНС:
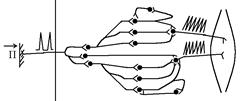 а) посилюється за рахунок поширення по дивергентним нейронним ланцюгам Ù проходить іррадіація збудження, його широке поширення. Збудження як би множиться – проходить його мультиплікація.
а) посилюється за рахунок поширення по дивергентним нейронним ланцюгам Ù проходить іррадіація збудження, його широке поширення. Збудження як би множиться – проходить його мультиплікація.
б) посилюється за рахунок того, що збудження, яке множилось в дивергентних ланцюгах, сходиться до тіл кількох еферентних нейронів по конвергентних ланцюгах – на тілах еферентних нейронів інтенсивно проходить сумація ЗПСП Ù висока ступінь збудження нейронів (велика частота ПД в РРН).
в) подовжується за рахунок включення в систему передачі інформації збуджуючих кільцевих нейронних ланцюгів.
Таким чином, процес іррадіації збудження по дивергентним нейронним ланцюгам є важливим механізмом посилення біологічно важливих аферентних сигналів в ЦНС. Спостерігати іррадіацію збудження можна на спінальній жабі, поступово посилюючи силу подразника, який діє на її шкіру (механічний – щипок пінцетом, хімічний – розчин кислоти). При малій силі подразника отримуємо відповідь у вигляді локального рефлексу – проходить згинання тільки тієї кінцівки, на шкіру якої діє подразник. При підвищенні сили подразника в реакцію у відповідь включаються все більше органів-ефекторів (скелетних м’язів). В кінцевому результаті спостерігається генералізована реакція у відповідь у вигляді координованого скорочення м’язів кінцівок і тулуба жаби Ù згинальні та розгинальні рухи кінцівок і тулуба.
Але, щоб в умовах іррадіації збудження проходили координовані рухи, необхідно обмеження поширення збудження в межах ЦНС – одні центри повинні збуджуватись, а інші – переходити в стан гальмування. Обмежує поширення збудження в ЦНС реципрокне (спряжене) гальмування – завдяки йому при іррадіації збудження в ЦНС одні центри збуджуються, а інші (антагоністичні) – гальмуються. Реципрокне гальмування являється дуже важливим механізмом координації процесів збудження та гальмування в ЦНС (відповідно, механізмом координації функцій організму). Якщо заблокувати ці процеси (реципрокного постсинаптичного гальмування) за допомогою стрихніну, тварина на самий легкий подразник відповідає генералізованою, але не координованою судомною реакцією – скороченням всіх скелетних м’язів.
Явище домінанти та його фізіологічне значення.
Принцип домінанти являється одним із основних принципів організації рефлекторної діяльності. Воно заключається в наявності в ЦНС домінантного центру збудження. Домінантний центр збудження характеризується слідуючими властивостями:
1. Високою збудливістю нейронів;
2. Інертністю (тривалістю) збудження – воно зберігається в цьому центрі протягом тривалого відрізку часу після закінчення дії подразника.
Такі властивості нейронів домінантного центру можуть бути обумовлені
двома механізмами:
1. Тривалим надходженням інформації про дію якого-небуть подразника по аферентним нервовим шляхам;
2. Тривалим підвищенням концентрації яких-небуть гормонів в плазмі крові.
Висока збудливість нейронів та інертність процесів збудження в домінантному центрі викликають такі особливості його функціонування:
а) він гальмує інші центри ЦНС по механізму одночасного спряженого гальмування;
б) завдяки високій збудливості нейронів і спряженому гальмуванню інших центрів, домінантний центр як би “притягує” до себе аферентні сигнали, що призначені для інших центрів, внаслідок чого ці центри загальмовані і не відповідають на сигнал. Доходячи до домінантного центру ці сигнали викликають збудження його нейронів Ù відповідна рефлекторна реакція, пов’язана з збудженням нейронів домінантного вогнища.
Як наслідок, рефлекторна відповідь, пов’язана з збудженням домінантного центру, може виникнути у відповідь на будь-яке подразнення!!! Наприклад, поглажування собаки з переповненим сечовим міхуром може викликати у неї рефлекс сечовипускання (стійка аферентація від механорецепторів сечового міхура Ù формування домінантного вогнища збудження в центрі рефлексу сечовипускання); весною у самця жаби дотик до шкіри викликає обіймальний рефлекс, що пояснюється високою концентрацією статевих гормонів Ù формування домінантного вогнища збудження в центрі обіймального рефлексу.
Тобто, характер рефлекторної відповіді залежить не тільки від виду діючого подразника, а також від стану нервових центрів, тобто, домінантний центр як би підпорядковує собі діяльність інших центрів.
Рухові рефлекси спинного мозку, їх рефлекторні дуги, фізіологічне значення.
Кажучи про роль різних рівнів ЦНС в регуляції рухових функцій, необхідно висвітлювати такі питання:
1. Особливості будови (сегментарні механізми, надсегментарні утворення).
2. Контроль за станом зовнішнього, внутрішнього середовища, рухового апарату (від яких рецепторів і про що надходить інформація).
3. Регуляція рухових функцій:
- тонічних, тобто регуляція тонусу м’язів; тонус скелетних м’язів – скорочення невеликої сили, але тривалі. Тонус забезпечує збереження пози – рівноважного положення тіла у просторі. Тонус найбільш виражений у м’язах тулуба і проксимальних відділах кінцівок.
- фазичних, тобто таких, при яких добре виражені фази скорочення і розслаблення; за рахунок таких скорочень відбувається переміщення тіла у просторі та рух кінцівок відносно тулуба.
Читайте також:
- Асинхронний генератор з конденсаторним збудженням.
- Аутогенне гальмування
- Біоелектричні явища і збудження в тканинах.
- Вибір елементів пуско-гальмівного реостата для двигунів послідовного збудження
- Вибір елементів пускового (гальмівного) реостата для двигунів змішаного збудження
- Вибір обмеження у режимі гальмування
- Види умовних рефлексів, механізм утворення та зберігання. Гальмування.
- Види центрального гальмування. Механізми розвитку пре- та постсинаптичного гальмування.
- Властивості м’язового волокна. Механізми утворення та передачі збудження, скорочення скелетних м’язів.
- Гальмування в ЦНС.
- Гальмування умовних рефлексів
- Гальмування умовних рефлексів
| <== попередня сторінка | | | наступна сторінка ==> |
| Види центрального гальмування. Механізми розвитку пре- та постсинаптичного гальмування. | | | Рухові рефлекси спинного мозку, їх рефлекторні дуги, фізіологічне значення. |
|
Не знайшли потрібну інформацію? Скористайтесь пошуком google: |
© studopedia.com.ua При використанні або копіюванні матеріалів пряме посилання на сайт обов'язкове. |