РЕЗОЛЮЦІЯ: Громадського обговорення навчальної програми статевого виховання
ЧОМУ ФОНД ОЛЕНИ ПІНЧУК І МОЗ УКРАЇНИ ПРОПАГУЮТЬ "СЕКСУАЛЬНІ УРОКИ"
ЕКЗИСТЕНЦІЙНО-ПСИХОЛОГІЧНІ ОСНОВИ ПОРУШЕННЯ СТАТЕВОЇ ІДЕНТИЧНОСТІ ПІДЛІТКІВ
Батьківський, громадянський рух в Україні закликає МОН зупинити тотальну сексуалізацію дітей і підлітків
Відкрите звернення Міністру освіти й науки України - Гриневич Лілії Михайлівні
Представництво українського жіноцтва в ООН: низький рівень культури спілкування в соціальних мережах
Гендерна антидискримінаційна експертиза може зробити нас моральними рабами
ЛІВИЙ МАРКСИЗМ У НОВИХ ПІДРУЧНИКАХ ДЛЯ ШКОЛЯРІВ
ВІДКРИТА ЗАЯВА на підтримку позиції Ганни Турчинової та права кожної людини на свободу думки, світогляду та вираження поглядів
- Гідрологія і Гідрометрія
- Господарське право
- Економіка будівництва
- Економіка природокористування
- Економічна теорія
- Земельне право
- Історія України
- Кримінально виконавче право
- Медична радіологія
- Методи аналізу
- Міжнародне приватне право
- Міжнародний маркетинг
- Основи екології
- Предмет Політологія
- Соціальне страхування
- Технічні засоби організації дорожнього руху
- Товарознавство продовольчих товарів
Тлумачний словник
Авто
Автоматизація
Архітектура
Астрономія
Аудит
Біологія
Будівництво
Бухгалтерія
Винахідництво
Виробництво
Військова справа
Генетика
Географія
Геологія
Господарство
Держава
Дім
Екологія
Економетрика
Економіка
Електроніка
Журналістика та ЗМІ
Зв'язок
Іноземні мови
Інформатика
Історія
Комп'ютери
Креслення
Кулінарія
Культура
Лексикологія
Література
Логіка
Маркетинг
Математика
Машинобудування
Медицина
Менеджмент
Метали і Зварювання
Механіка
Мистецтво
Музика
Населення
Освіта
Охорона безпеки життя
Охорона Праці
Педагогіка
Політика
Право
Програмування
Промисловість
Психологія
Радіо
Регилия
Соціологія
Спорт
Стандартизація
Технології
Торгівля
Туризм
Фізика
Фізіологія
Філософія
Фінанси
Хімія
Юриспунденкция
РОЗПАД мРНК
Полінуклеотидфосфорилаза
РНК-вмісні віруси
Нарешті, реовируси містять в якості генома близько 10 різних дволанцюгових молекул РНК, які можна позначити як (±) –РНК.
Принцип репродукції цих вірусів такий же, якби геномом була дволанцюгова ДНК. Тільки ключовим ферментом є РНК-синтетаза.
Неодноразово транскрибуючі (-) -ланцюги (±) РНК, цей фермент утворює (+) -ланцюги, які виступають в якості мРНК. А для накопичення нових дволанцюгових молекул (±) -РНК, очевидно, потрібне використання як матриці обох ланцюгів вірусної РНК.
У деяких клітинах (як бактерійного, так і тваринного походження) є фермент полінуклеотидфосфорилаза (ПНФаза). Він каталізує реакцію, яка може йти як в прямому, так і в зворотному напрямах.
Прямий напрямок реакції - розщеплення РНК шляхом фосфоролізу. Це означає, що як літичного агента використовується не вода, а фосфорна кислота. Завдяки цьому продуктами реакції є не нуклеозидмоно-, а нуклеозиддифосфати (рНДФ).
Очевидно, суть зворотної реакції - об'єднання рНДФ в полінуклеотидний ланцюг (з вивільненням фосфатних залишків) :
вільні рНМФ → залишки рНДФ в полинуклеотидной ланцюзі + фосфат
При цьому матриця не використовується, і тому склад і довжина продукту визначаються випадковими обставинами - концентрацією вільних рНДФ в середовищі і тому подібне.
Невідомо, чи грає ця реакція яку-небудь роль в клітині.
У експерименті ж ПНФаза довгий час широко використовувалась для синтезу полірибонуклеотидів необхідного складу, які потім служили як мРНК для розшифрування генетического коду.
Наприклад, синтезувавши пів(У), виявили, що в білок-синтезуючій системі цей полінуклеотид викликає утворення пептиду з одних залишків фенілаланіна. Звідси - висновок про те, що триплет УУУ кодон фенілаланіна.
Коли ж був синтезований полімер пів(УГ) із закономерним чергуванням У і Г, тобто що містить триплети двох видів (УГУ і ГУГ), виявилось, що під його впливом в пептид з тією ж черговістю включаються дві амінокислоти - цистеїн і валін. Отже, кодон цистеїна - УГУ, а кодон валіну - ГУГ.
І так далі: так само був розшифрований зміст усіх 64 кодонів.
Досі ми говорили про утворення РНК. Тепер же коротко обговоримо, як розпадаються РНК; найцікавіше це питання відносно матричних РНК.
Правда, на відміну від розглянутих вище процесів, розпад мРНК відбувається (у еукаріот) не в ядрі клітини, а в цитоплазмі - там, де вони функціонують. Проте зручніше звернутися до цієї проблеми зараз щоб висвітлити відразу і другу сторону обміну мРНК.
Руйнування мРНК бактерій з 5´ -конца-ефект положення
У бактерій (принаймні, у деяких їх видів) розпад мРНК може починатися з 5'- кінця, тобто відбуватися в тому ж напрямі (5 '→3'), в якому мРНК синтезується на ДНК і за тим транслюється рибосомами.
Нагадаємо також, що у бактерій немає ядра, а мРНК виявляються поліцистронними, тобто досить довгими.
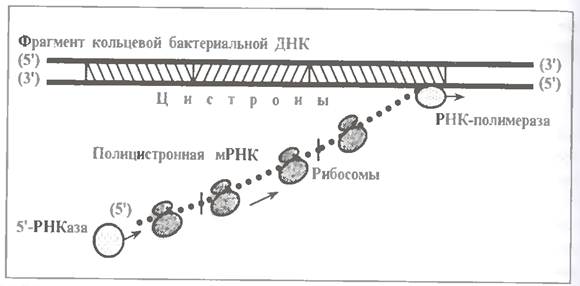
Усі ці причини призводять до того, що в якісь моменти часу один і той же ланцюг мРНК може одночасно брати участь в трьох процесах (див.мал.) :
· своєму утворенні на ДНК (в ході чого продовжує подовжуватись зі свого 3'-кінця);
· своєму функціонуванні, тобто поцистронній трансляції рибосомами;
· своєму розпаді, тобто поступовому укороченні з 5 '- кінця за рахунок дії 5'-РНКази.
У свою чергу, цей феномен може призводити до т. з. ефекту положення. Суть його в тому, що кратність трансляції будь-якого цистрона (тобто кількість разів, скільки він устигає прочитуватися рибосомами) залежить від положення цього цистрона в опероні. У одних випадках при переході від першого цистрона до останнього кратність трансляції закономірно зменшується, а в інших так само закономірно збільшується.
Дійсно, допустимо, що 5'-РНКаза зв'язалася з 5'- кінцем мРНК після того, як з ним встигли зв'язатися 10 рибосом: така кратність трансляції першого цистрону.
І нехай 5'-РНКаза укорочує мРНК з 5 '- кінця швидше, ніж РНК-полімераза подовжує мРНК із 3'-конца.
Тоді кожен наступний цистрон існуватиме менший час, чим попередній. Відповідно, менша кількість рибосом встигне зв'язатися з ним і здійснити трансляцію. Тобто в цьому прикладі кратність трансляції буде в опероні закономірно зменшуватися.
Кокретні ж значення тривалості життя цистронів мРНК у бактерій дуже невеликі: зазвичай варіюють в межах 2 3 хвилин.
Читайте також:
- Активність. Закон радіоактивного розпаду
- Антитоталітарні демократичні революції у країнах Східної Європи. Розпад Югославії. Об’єднання Німеччини
- Вплив ліганда білку на розпад мРНК
- Вплив продуктів трансляції на розпад мРНК
- Еволюція господарських форм в країнах Західної Європи на етапі розпаду натурального господарства
- Економічний розвиток і економічна думка східних та західних українських земель у період розпаду феодального господарства
- Електролітична дисоціація (йонізація) — це розпад електролітів на сольватовані (гідратовані) йони під дією молекул розчинника.
- Лекція 16. «Перебудова» і розпад СРСР. На шляху до незалежності
- Питання 2. Розпад глікогену
- Поняття радіоактивності та закон радіоактивного розпаду
- Поступовий занепад і розпад Київської Русі, утворення самостійних князівств.
- Причини розпаду СРСР.
| <== попередня сторінка | | | наступна сторінка ==> |
| РНК-вмісні віруси | | | Роль АУ-элементів |
|
Не знайшли потрібну інформацію? Скористайтесь пошуком google: |
© studopedia.com.ua При використанні або копіюванні матеріалів пряме посилання на сайт обов'язкове. |