РЕЗОЛЮЦІЯ: Громадського обговорення навчальної програми статевого виховання
ЧОМУ ФОНД ОЛЕНИ ПІНЧУК І МОЗ УКРАЇНИ ПРОПАГУЮТЬ "СЕКСУАЛЬНІ УРОКИ"
ЕКЗИСТЕНЦІЙНО-ПСИХОЛОГІЧНІ ОСНОВИ ПОРУШЕННЯ СТАТЕВОЇ ІДЕНТИЧНОСТІ ПІДЛІТКІВ
Батьківський, громадянський рух в Україні закликає МОН зупинити тотальну сексуалізацію дітей і підлітків
Відкрите звернення Міністру освіти й науки України - Гриневич Лілії Михайлівні
Представництво українського жіноцтва в ООН: низький рівень культури спілкування в соціальних мережах
Гендерна антидискримінаційна експертиза може зробити нас моральними рабами
ЛІВИЙ МАРКСИЗМ У НОВИХ ПІДРУЧНИКАХ ДЛЯ ШКОЛЯРІВ
ВІДКРИТА ЗАЯВА на підтримку позиції Ганни Турчинової та права кожної людини на свободу думки, світогляду та вираження поглядів
- Гідрологія і Гідрометрія
- Господарське право
- Економіка будівництва
- Економіка природокористування
- Економічна теорія
- Земельне право
- Історія України
- Кримінально виконавче право
- Медична радіологія
- Методи аналізу
- Міжнародне приватне право
- Міжнародний маркетинг
- Основи екології
- Предмет Політологія
- Соціальне страхування
- Технічні засоби організації дорожнього руху
- Товарознавство продовольчих товарів
Тлумачний словник
Авто
Автоматизація
Архітектура
Астрономія
Аудит
Біологія
Будівництво
Бухгалтерія
Винахідництво
Виробництво
Військова справа
Генетика
Географія
Геологія
Господарство
Держава
Дім
Екологія
Економетрика
Економіка
Електроніка
Журналістика та ЗМІ
Зв'язок
Іноземні мови
Інформатика
Історія
Комп'ютери
Креслення
Кулінарія
Культура
Лексикологія
Література
Логіка
Маркетинг
Математика
Машинобудування
Медицина
Менеджмент
Метали і Зварювання
Механіка
Мистецтво
Музика
Населення
Освіта
Охорона безпеки життя
Охорона Праці
Педагогіка
Політика
Право
Програмування
Промисловість
Психологія
Радіо
Регилия
Соціологія
Спорт
Стандартизація
Технології
Торгівля
Туризм
Фізика
Фізіологія
Філософія
Фінанси
Хімія
Юриспунденкция
ЗАГАЛЬНЕ УЯВЛЕННЯ ПРО ЧИННИКИ
Сучасне розповсюдження організмів визначається в насамперед умовами середовища, в якому вони мешкають. Середовище впливає на організми різними складовими її елементами. Кожен елемент середовища є чинником (діючим фактором), оскільки він тією чи іншою мірою впливає на ті або інші організми. Якщо раніше вважали, що є елементи середовища, що не впливають на організми, то тепер уже достатньою мірою з'ясовано, що кожен елемент середовища діє хоча б на деякі організми. Середовище складається з чинників, які повинні бути класифіковані. Перш за все, виділяють чинники абіотичні (неживої природи) і чинники біотичні (живої природи). Особливо виділяють чинники антропічні (вплив людини). До абіотичних чинників належать в першу чергу кліматичні чинники, що впливають на всі організми, що мешкають в тій або іншій місцевості, — це вологість повітря і кількість опадів, температура повітря, кількість світла, що падає на землю, і фотоперіодичність, тобто співвідношення тривалості дня і ночі, а також вітровий режим, тиск повітря, газовий склад атмосфери і в першу чергу вміст СО2.
Другу групу абіотичних чинників для наземних організмів складають чинники едафічні, тобто ґрунтові (хімічні, фізичні і механічні особливості ґрунтів і підґрунтя). Третя група чинників, що визначає існування водних організмів, ‑ гідрологічні, ‑ зміни прозорості та освітленості залежно від глибини і каламутності водної товщі, зміни тиску з глибиною тощо.
Четверту групу абіотичних чинників складають чинники орографічні, особливості рельєфу. Вони мають як місцевий вплив (вплив експозиції і крутизни схилу, висота над рівнем моря), так і віддалений вплив, непрямий (наприклад, вплив того або іншого гірського хребта на розподіл кліматичних чинників, перш за все вологості й температур).
До біотичних чинників відносяться різноманітні впливи організмів один на одного, різні за своїм характером ‑ симбіотичні, антагоністичні, конкурентні та інші. Це взаємостосунки між організмами, що відносяться до різних систематичних груп і до одного і того ж виду.
Раніше до біотичних чинників відносили і дію людини на організми, проте нині вважають за краще виділяти людську діяльність в особливу категорію чинників, яку правильніше називати чинниками антропічними (іноді вживають термін — антропогенні). Отже, система чинників виглядає так:
1. Чинники абіотичні (кліматичні, гідрологічні, едафічні, орографічні).
2. Чинники біотичні.
3. Чинники антропічні.
Кожен чинник має певну амплітуду значень. Це стосується і температур, і вологості, й інших чинників. Для різних організмів набір значень чинника, при якому він може існувати, неоднаковий. Найменше значення чинника, при якому може існувати той або інший організм, називається його мінімумом, найбільше ‑ максимумом. Життя будь-якого організму протікає між мінімальним і максимальним значеннями даного чинника. Найбільш сприятливе для життя організму значення чинника є його оптимумом, або оптимальним значенням. Як правило, на шкалі значень чинника оптимум дещо зміщений у бік максимуму. Ті значення чинника, при яких організм існує, але тією чи іншою мірою пригноблений, називаються його песимальними значеннями (песимумом). Таким чином, песимум лежить між оптимумом і мінімумом, між оптимумом і максимумом. До деяких чинників поняття оптимуму непридатне. Так, всі значення іонізуючої радіації шкідливі для організму. Чим вони вище, тим гірше для організму. До цього слід додати, що виконання організмом одних функцій вимагає одного оптимуму значень даного чинника, інших ‑ іншого, те саме стосується значень мінімуму і максимуму. Так, яйця аскариди ‑ одного з паразитів людини ‑ можуть розвиватися від +13 до +36° при оптимумі 30°, а зберігати життєздатність можуть при температурах від ‑ 20 до +38°, іншими словами, вони від ‑ 20 до +13° і від +36 до +38° хоч і зберігають життєздатність, але розвиватися не можуть.
Організми, що можуть існувати при широкій амплітуді чинників, називаються еврибіонтними, а організми, для яких існування можливо лише при вузькій амплітуді чинників, іменуються стенобіонтними. Природно, що еврибіонтні організми можуть за інших рівних умов займати більші області, ніж організми стенобіонтні. Такий самий поділ організмів можливий і стосовно до будь-якого окремого чинника: організми можуть бути еври- і стенотермними (по відношенню до температур середовища), еври- і стеногігробіонтними (по відношенню до умов вологості), еври- і стеногалинні (по відношенню до солоності середовища) тощо. Іншими словами, ми можемо говорити про організми, що мають широку і вузьку екологічну амплітуду по відношенню до всіх або до одного з чинників.
Німецький агрохімік Ю. Лібіх встановив правило (іноді його називають законом), згідно з яким існування організму визначається тим чинником, який перебуває в мінімумі. У. П. Гептнер застосував це правило в зоогеографії: нормальне і тривале існування виду в якій-небудь місцевості неможливе, якщо хоч би один чинник розвинений в мірі, що перебуває нижче за мінімум потреб виду. Проте насправді чинники діють не ізольовано один від одного, а спільно і тому можуть тією чи іншою мірою заміщувати один одного, як вказав швейцарський учений Е. Рюбель. Прикладів таких заміщуючих дій чинників дуже багато. Так, всі ми знаємо, що багато птахів залишають широти помірного поясу восени, відлітаючи на південь, де тепліше. Однак якщо птахи отримують в достатку корм, то вони можуть зимувати і там, де вони зазвичай взимку відсутні. Рослини, які не можуть жити під покривом лісу в помірному поясі через нестачу світла, в тропічному лісі, при вищих температурах, переносять значно більшу нестачу освітлення. У першому випадку багата їжа «замістила» знижені температури, в другому — високі температури «замістили» нестачу світла.
Розглянемо дію різних чинників та їх зміни в угрупованнях рослин і тварин.
КЛІМАТИЧНІ ЧИННИКИ
Світло є одним з основних кліматичних чинників, що визначають можливості існування організмів. Тільки на світлі відбувається фотосинтез і, отже, можуть існувати фотосинтезуючі рослини. Нерухомим рослинам доводиться пристосовуватися до певного рівня освітлення. Вони можуть бути поділені по відношенню до світла на три групи: світлолюбні, такі, що мешкають в умовах доброї освітленості, тіньолюбні, такі, що мешкають завжди в умовах значної затіненості, і тіньовитривалі, що надають перевагу життю на світлі, але можуть існувати і в тіні. Рухомі тварини, крім активного вибору місцепроживання з певними умовами освітленості, мають можливість будувати свою добову активність так, щоб отримувати певну кількість світла. Серед них розрізняють тварин денних, нічних і присмеркових. Будучи активними лише певну частину доби, решту часу тварини проводять в норах і притулках, ховаються під покривом листя, серед лісової підстилки тощо. Характер активності тварин може змінюватись залежно від пори року, метеорологічної обстановки і умов освітлення. Так, ховрахи, що мешкають в напівпустелях, в жаркі дні бувають активними у ранкові і вечірні години, а полуденний час проводять в норах; у похмурі дні вони можуть з'являтися на поверхні й вдень. Багато птахів пустелі, що уникають в жаркі літні дні з'являтися на відкритих місцях, восени, взимку і весною виявляють максимум активності в денні години. Полівки в затінених ділянках виявляють цілодобову активність з максимумом вночі, а на освітлених ділянках активні лише вночі.
Рослини, що виросли в тіні, характеризуються тонким, ніжним листям з|із| відносно слабо розвиненою палісадною|живопліт| тканиною і добре розвиненою губчастою. У|в,біля| цих рослин міжвузля|міжвузля| стебел|стеблин| тонкі, витягнуті, механічна тканина розвинена слабо. У|в,біля| рослин, що виросли на світлі, листя щільні з|із| добре розвиненою палісадною|живопліт| тканиною, стебла|стеблини| товсті, з|із| короткими міжвузлями|. Під дією світла рослини через зміни швидкості процесів росту можуть повертатися|обертатися| у бік джерела світла (позитивний фототропізм), убік, протилежний до джерела світла (негативний|заперечний| фототропізм), і перпендикулярно до напряму|направленню| світлових променів (трансверсальний фототропізм). Прикладом|зразком| останнього є|з'являється,являється| виникнення так званої листової мозаїки, коли листя розташовується так, щоб найкраще вловлювати сонячне проміння. Навпаки, в тих випадках, коли світла багато, листя може звисати з дерева, так що сонячні промені ковзають вздовж їх поверхні (наприклад, у|в,біля| австралійського евкаліпта) або обернені ребрами на північ і на південь, а площинами|плоскістю| на захід і схід (у|в,біля| компасних рослин аридних| районів світу).
Якщо нерухомі рослини реагують на світло за допомогою зміни свого положення|становища| по відношенню до джерела світла — фототропізму, то рухомі|жваві,рухливі| тварини володіють фототаксисом — вони здатні|здібні| рухатися|сунутися| у напрямку до світла (позитивний фототаксис) або від світла (негативний|заперечний| фототаксис).
Частина світла, падаючого на поверхню рослин, що створюють угруповання, відбивається, частина|частка| поглинається рослинами, а частина|частка| проникає всередину угруповання і там або поглинається рослинами нижніх ярусів, або досягає поверхні ґрунту. Природно, що при цьому сила світла поступово знижується, а його спектральний склад змінюється. Отже, рослинні |спілки|угруповання диференціюють світлові умови, характерні|вдача| для якої-небудь місцевості: неоднакові умови освітленості створюються в різних угрупованнях|спілках|, на різних ділянках одного і того самого угруповання|спілки|, в одному і тому самому угрупованні в різні сезони року. Кількість світла, проникаючого|проникного| під запону дерев верхнього ярусу, пов'язана з відносним мінімумом світла, тобто з|із| тією його величиною, при якій рослина даного виду ще може асимілювати. Для модрини європейської відносний мінімум сили світла складає в середньому близько 20% від повного|цілковитого| денного освітлення, для сосни звичайної — близько 10, для дуба черешкового — 3,8, для ялини європейської — 3,0, для бука лісового — 1,5, для самшиту — 1%. Це означає|значить|, що в модриновому лісі значно світліше, ніж в сосновому, а в сосновому світліше, ніж в ялиновому, дубовому і тим більше буковому і самшитовому. Останній є|з'являється,являється| найменш світлопроникним з|із| лісів, що зростають у помірних широтах та субтропіках. Ще темніше під покривом вологого|вогкого| тропічного лісу, куди проникає лише 0,2% повного|цілковитого| денного світла. Для лісових трав’янистих рослин освітленість значну частину|частку| дня може бути нижча за так звану компенсаційну величину, тобто такої освітленості, при якій асиміляція і дихання врівноважують|зрівноважують,урівноважують| один одного (нижче за цю величину рослина у принципі|в принципі| існувати не може, оскільки|тому що| його дихання стає інтенсивнішим, ніж асиміляція). Виявляється, що така тіньова рослина, як кислиця, може вдовольнятися короткими проміжками часу, коли освітленість перевищує компенсаційну величину завдяки падінню на рослину сонячного відблиску.
У лісах, утворених деревами з|із| опадаючим листям, навесні освітленість поверхні ґрунту велика, і в цей час в лісі зацвітають деякі світлолюбні трави. Під покривом хвойного або вічнозеленого лісу сезонні зміни освітленості незначні.
Крім загальної|спільної| кількості світла, що його отримують організми, для них має велике значення і фотоперіодизм, тобто характер|вдача| чергування світлих і темних періодів (дня і ночі). Фотоперіодизм змінюється від приекваторіальних| широт до помірних і холодних поясів. Поблизу|неподалік| екватора тривалість дня і ночі мало розрізняється протягом круглого року; у високих широтах влітку день буває дуже тривалим, а ніч коротка, взимку — навпаки. Мешканці|пожильці| цих широт — організми тривалого дня, оскільки влітку, в період їх активності, день значно перевищує ніч; мешканці|пожильці| тропіків — організми короткого дня. Мешканцям|пожильцям| різних широт властиві пристосування до певного фотоперіодизму.
Так, рослини тривалого дня потребують високої тривалості світлого періоду, щоб накопичити|скупчити,нагромадити| достатню кількість органічної речовини. Якщо їх помістити в умови короткого дня, вони будуть успішно вегетувати, але|та| не зможуть перейти до утворення генеративних органів, оскільки|тому що| світлої частини|частці| доби недостатньо для накопичення достатніх запасів органічної речовини. Рослини короткого дня, перенесені в умови тривалого дня, не зможуть перейти від вегетативного розвитку до генеративного. Це пов'язано з тим, що вони на батьківщині протягом тривалішої ночі встигають|устигають| звільнити|визволити| листя від накопичених органічних речовин і тим самим підготувати асиміляційний апарат для успішного фотосинтезу до початку наступного|такого| дня, а в низьких широтах ніч надто коротка для цього. У|в,біля| тварин помірного поясу (тварин тривалого дня) осіннє скорочення дня веде до припинення процесів розмноження. Весняне подовження|видовження| дня відновлює|поновлює| здатність до розмноження. Механізм дії цього подовження|видовження|, мабуть|певне|, такий: подовження|видовження| дня діє на гіпофіз — придаток головного мозку, той стимулює роботу ендокринних органів, які передають сигнал, що надійшов|вчинив|, статевим залозам. Збільшення тривалості світлового дня шляхом додаткового щодобового освітлення пташників дозволяє підвищити яйценосність свійської птиці.
З іншого боку, врожайність яблуні — дерева короткого дня — може бути підвищена, якщо за умов тривалого дня закривати|зачиняти| її крону декілька годин на добу дерев'яним непроникним для світла шатром|наметом|. Це одночасно призводить також до підвищення морозостійкості цього південного дерева, що може служити прикладом|зразком| заміщення дії чинників|факторів|, про яку ми говорили раніше.
Одним з найважливіших|поважних| кліматичних чинників|факторів|, що визначають розміщення організмів по земній кулі, є|з'являється,являється| температур а. Ряд|лава,низка| тварин може виробляти постійну внутрішню температуру тіла. Такі тваринні називаються гомеотермними|, на відміну від тварин пойкілотермних, температура тіла яких непостійна і визначається температурою середовища|середи|. Проміжну групу складають тварини гетеротермні|, які в активному стані гомеотермні|, а при впаданні в тривалу сплячку, що триває іноді|інколи| до напівроку і більш, втрачають|розгублюють| здібність до терморегуляції. У цей період число вдихів і видихів у|в,біля| них складає 1-2 на хвилину, з|із| такою самою швидкістю скорочується серце.
Абсолютної межі між пойкілотермними і гомеотермними| тваринами не існує. Багато пойкілотермних тварин можуть при русі продукувати енергію, що призводить до підйому температури їх тіла іноді|інколи| значно вище за температуру середовища|середи|. Так, температура тіла багатьох комах (джмелі, великі метелики) під час їх польоту може перевищувати температуру середовища|середи| на 10-15°. Так само і деякі пойкілотермні водні тварини в змозі|спроможний| продукувати значні кількості тепла, незважаючи на високу швидкість тепловіддачі у водному середовищі|середі|. Наприклад, риба тунець при швидкому русі може мати температуру тіла, що перевищує на 8-9° температури води.
З іншого боку, ті пташенята, які виходять з|із| яйця нездібними до самостійного життя, мають не цілком|сповна| постійну температуру тіла: вона падає, коли батьки не гріють їх своїм тілом, і підіймається|підіймається|, коли батьки знаходяться|перебувають| на гнізді. Температура сліпих голих дитинчат звичайної полівки може падати до 8-9 і навіть 4-5°, а при зігріванні звірят, що остигнули|вичахнули|, знов|знову,щойно| підіймається|підіймається| до нормальної для них температури 28-30°. Поява здібності до терморегуляції збігається у|в,біля| дитинчат з|із| їх прозрінням.
Температура тіла дорослих полівок і деяких інших дрібних|мілких| гризунів коливається|вагається| в межах 28-41° залежно від умов, в яких звір мешкає, і від стану звірок (так, звірок з|із| намоклою шкіркою швидко охолоджується). При пониженні температури тіла нижче 28° звірок швидко гине. Тому для них безпечні літні дощі, які не викликають|спричиняють| переохолодження тварин, але|та| при осінніх дощах звірам загрожує загибель в результаті|унаслідок,внаслідок| пониження температури тіла нижче за мінімальну. Коливання температури тіла відмічені також для колібрі, стрижів і деяких інших птахів|птиць|.
Гомеотермність («теплокровність|») — істотна|суттєва| перевага тварини в боротьбі за існування, саме тому гомеотермні| тварини — ссавці і птахи|птиці| — в порівнянні з пойкілотермними найширше розселені по Землі|ґрунту|.
Всі організми, що населяють нашу планету, по відношенню до температури середовища|середи| можуть бути евритермними (що мешкають при значних коливаннях цього чинника|фактору|) і стенотермними (що мешкають при вузьких коливаннях температур). Деякі стенотермні організми мешкають тільки|лише| при високих температурах. Це теплолюбні|теплолюбні| форми. Частина|частка| з|із| них може мешкати в теплих джерелах з|із| температурою 70-80°. У таких умовах живуть представники синьо-зелених водоростей і бактерій. Ряд|лава,низка| тварин і деякі водорості мешкають в джерелах з|із| температурою 50-60°. Зазвичай білки згортаються при 40-45°. Тому незрозуміло, які особливості дозволяють цим організмам виживати при таких температурах. Інші стенотермні організми є|з'являються,являються| холодолюбними. Стенотермними можуть бути не тільки|не лише| пойкілотермні, але і гомеотермні| тварини. Так, мешканці|пожильці| тропічних широт — носороги, слони, напівмавпи, велика частина|частка| мавп, папуги — теплолюбні|теплолюбні|; білий ведмідь, пінгвіни і інші мешканці|пожильці| полярних країн — холодолюбні.
Температурний режим на суші змінюється в двох напрямах|направленнях|: середні річні температури повітря знижуються від тропіків до полярних широт, амплітуди добових і річних температур зростають від околиць в глиб континентів. Будь-яка точка земної кулі характеризується певними середніми добовими, місячними і річними амплітудами температур, певною тривалістю і температурним режимом різних пір року. Ці особливості температурного режиму обмежують можливості|спроможності| існування організмів в тому або іншому місці.
Рослинні угруповання здійснюють істотний|суттєвий| вплив на річний і добовий хід температур. Температура ґрунту на глибині 10 см має найбільш рівний хід і дещо нижча під непорушеним лучним угрупованням |спілкою|, дещо вища і характеризується більшими коливаннями під лугом з|із| видаленим|віддаленим| моховим покривом|покривалом|. Врешті, вищі температури з|із| помітними коливаннями властиві ділянкам лугу, на яких був видалений|віддалений| і моховий, і трав'яний покрив|покривало|. Ще сильніше впливають на температурний режим ліси. Під запоною лісу зменшуються добові й сезонні амплітуди температур, а середні добові і річні температури нижчі, ніж на відкритому|відчиненому| місці.
Розвиток рослин і пойкілотермних тварин перебуває в прямій залежності від температури середовища|середи|. Він починається з тієї температури, яка для даного виду є|з'являється,являється| біологічним нулем (нижче за яку він розвиватися не може). Далі, до досягнення оптимальної температури підвищення температури прискорює розвиток. Тому в екології прийнято обчислювати|обчисляти,вичисляти| суми ефективних температур, необхідних для проходження тих або інших стадій розвитку.
Звичайний|звичний| метод підрахунку кількості тепла, необхідного для життєдіяльності організму або для виконання їм яких-небудь функцій, — метод підрахунку суми ефективних температур. За нижню точку відліку беруть звичайно той мінімум, при якому починається|розпочинається,зачинається| розвиток організму, або який-небудь умовний рубіж (0° +5°, + 10°). Знаючи на основі дослідів, яка сума тепла необхідна для розвитку організму (наприклад, для дозрівання плодів, зацвітання, перетворення личинки в лялечку, вильоту з|із| лялечки дорослої комахи тощо), визначаємо за формулою, скільки днів необхідно для виконання цієї функції 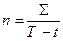
де п ‑ шукане число днів, Σ|β,α³λ| - сума тепла, необхідна для розвитку даного організму або виконання даної функції, Т — постійна температура періоду розвитку даного організму, t — нижній температурний поріг розвитку або вибраний нами умовний температурний рубіж. Якщо температура досліджуваного нами періоду непостійна, то просто складають середні добові температури доти, доки одержана|отримана| сума не досягне суми тепла, необхідного для розвитку даного організму або для виконання даної функції. День, коли це трапилося, і буде останнім з числа необхідних днів.
Проте|однак| визначення суми ефективних температур дає лише загальне|спільне| уявлення про терміни настання|наступу| того або іншого явища в житті рослини або тварини: коливання температур в період розвитку, перехідні за нижній поріг розвитку, можуть помітно порушити вказану вище картину. У цих випадках доводиться вносити поправки.
При оцінці впливу температурного режиму різних регіонів на гомеотермних| тварин застосовують два правила — Бергмана і Аллена. Не слід тільки|лише| абсолютизувати ці правила. Застосування|вживання| цих правил засноване на тому, що гомеотермні| тварини проводять|виробляють,справляють| тепло всією масою свого тіла, і віддають його своєю поверхнею. При збільшенні розмірів тварини відношення|ставлення| величини об'єму|обсягу| до поверхні стає в цьому відношенні сприятливішим. У цьому нас може переконати простий приклад|зразок|. Кубик з|із| довжиною ребра в 1 см має об’єм|обсяг| 1 см3 і поверхню 6 см2. Якщо довжину ребра збільшити удвічі|вдвічі| (до 2 см), то об’єм|обсяг| такого кубика буде дорівнювати 8 см3, а поверхня — 24 см2. Отже, при збільшенні лінійних розмірів удвічі|вдвічі| відношення|ставлення| об'єму|обсягу| до поверхні зміниться з 6 до 3, тобто стане сприятливішим з погляду співвідношення між виробництвом тепла і його віддачею. З|із| цього випливає, що в холодніших районах для тварини того самого виду сприятливіші великі розміри. Правило Бергмана й говорить про те, що в північніших районах нашої півкулі розміри тварин того самого виду стають більшими. Істотним|суттєвим| доповненням до цього правила служить правило Аллена, згідно з яким у|в,біля| тварин того самого виду, що мешкають в північніших районах, розміри частин|часток| тіла, що виступають поза |із|загальним|спільного| абрисом (вуха, хвіст, лапи) стають меншими.
Не менший вплив, ніж температура, чинить на загальні|спільні| риси|межі| розподілу організмів вологість|вогкість|. Є|наявний| організми евригігробіонтні|, що можуть існувати при самих різних значеннях вологості|вогкості|: ксерофільні — що мешкають в посушливих місцепроживаннях, мезофільні — в умовах середнього зволоження, гігрофильні — при високих значеннях вологості|вогкості|. При цьому способи пристосування організмів до життя в одних і тих самих умовах зволоження різноманітні|всілякі|. Так, за умовах значної сухості клімату одні рослини ніби «втікають|втікають|» від посухи, закінчуючи розвиток в той період, коли посуха ще не відчувається. Однорічні рослини-ефемери встигають|устигають| розвинутися з|із| насінини, вирости, відквітнути і принести плоди, а потім відмерти за той нетривалий період, коли в аридних| регіонах волога (снігова або дощова) ще значна. Багаторічні|багатолітні| рослини-ефемероїди, що розвиваються з|із| цибулин, бульб або кореневищ, за той самий нетривалий період приносять насіння, але|та|, на відміну від ефемерів, не відмирають повністю, а зберігають на період посухи підземні запасаючі органи — бульби, цибулини і кореневища.
Інші види, переважно водорості, лишайники і мохи, що мешкають в посушливих умовах — пойкілоксерофіти|, — висихають на період посухи так, що легко кришаться|, будучи взятими у руки, але|та| при зволоженні знов|знову,щойно| оживають і продовжують асимілювати. Деякі рослини запасають вологу на сухий період. Це сукуленти. Вони накопичують|скупчують,нагромаджують| вологу в стеблах|стеблинах| (стеблові|стеблисті| сукуленти) або в листі (листові сукуленти). Прикладами|зразками| перших можуть служити кактуси, кактусоподібні молочаї; інших – алое, агави, очитки тощо. Соковиті стебла|стеблини| і листя сукулентів погано переносять морози, тому вони зустрічаються переважно там, де холодного періоду немає або він короткочасний|. Ці рослини вкрай|надто| поволі|повільно| витрачають вологу: у|в,біля| них мало устечок, які при нестачі вологи дуже щільно закриваються|зачиняються|. Для фотосинтезу цим рослинам доводиться використовувати разом з|поряд з,поряд із| вуглекислотою повітря і той вуглекислий газ, який накопичується в міжклітинниках цих рослин при диханні. Тому ростуть|зростають| ці рослини дуже повільно. Зате вологу вони зберігають настільки надійно, що зірваний сукулент залишається в сухій кімнаті живим|жвавим| протягом декількох років.
Деякі рослини аридних| районів мають дуже довге коріння, яке може досягати ґрунтових вод навіть на глибині 16-18 м. Такі рослини, живучи в пустелі, знаходяться|перебувають| в умовах достатнього водопостачання, не відчуваючи нестачі вологи.
Інші рослини аридних| районів різко скорочують транспірування, маючи згорнуте в трубку|люльку| вузьке або дрібнорозсічене листя, покрите густим опушуванням, з|із| малим числом устечок|. Ці види не є|з'являються,являються| сукулентами, оскільки|тому що| у|в,біля| них немає соковитих, м'ясистих стебел|стеблин| або листя, але|та| в пристосуванні до життя в сухих регіонах вони йдуть тим самим шляхом|колії,дорозі| скорочення обміну з|із| середовищем|середою| і економії вологи. Проте|однак| завдяки обмеженості зв'язків з|із| середовищем|середою| ці рослини дуже повільно розвиваються.
Справжні ксерофіти| мають густу мережу|сіть| жилок і значне число устечок|. Клітинні|кліткові| стінки листових покривів|покривал| щільні, часто просочені восковою речовиною (кутинізовані|) або одягнені волосками. Випаровування крізь клітинні|кліткові| стінки, окрім устечок|, у|в,біля| цих рослин зведене до мінімуму, а устечка мають| здатність|здібністю| швидко закриватися|зачинятися| при нестачі вологи і так|настільки| само швидко відкриватися|відчинятися| при її достатній кількості. Добре розвинені механічні тканини не дозволяють їм зав'ядати|в'янути| при втраті вологи. Плазма цих рослин добре переносить тривале зневоднення. Таким чином, ці рослини пристосовані не стільки до нестачі вологи, скільки до її частих коливань.
Такі|настільки| ж різноманітні|всілякі| й пристосування тварин до життя в аридних| регіонах.
Деякі тварини запасають вологу. У сечовому міхурі|пузирі,пухирі| запасають воду черепахи Галапагоських островів і одна з австралійських пустинних жаб. Остання в період дощів роздувається, як куля, а до кінця сухого періоду, витративши воду, робиться|чиниться| плоскою.
Інші тварини, наприклад багато видів гризунів і комах, задовольняються тими незначними кількостями вологи, яку вони отримують з|із| їжею. Цю їжу складають рослини, особливо їх вологіші|вогкіші| підземні органи, а також деякі безхребетні тварини, що становлять їжу гризунів.
Треті впадають в літню сплячку, накопичивши|скупчивши,нагромадивши| перед цим значні кількості жиру. Під час сплячки, що проводять ці тварини в глибоких норах (такі, наприклад, ховрахи), процеси їх життєдіяльності різко уповільнені — серце робить|чинить| 1-2 ударів на хвилину, число вдихів і видихів також складає 1-2, тому запасів жиру, що розпадається під час сплячки на воду і вуглекислий газ, вистачає звіркам для підтримання життя.
Четверті накопичують|скупчують,нагромаджують| значні запаси жиру і витрачають його, не впадаючи в сплячку, довго|подовгу| не отримуючи ні їжі, ні пиття|питва|. Так, за рахунок метаболічного розпаду жирів і виділення в цьому процесі води компенсують нестачу вологи верблюди.
Види, що швидко бігають (антилопи) або літають (рябки|), можуть відвідувати водоймища, що знаходяться|перебувають| на значній відстані від місця їх проживання. Іноді|інколи| вони повертаються з водоймищ із|із| зобами, повними|цілковитими| води (рябки|).
Врешті, багато безхребетних весь посушливий період проводять у вигляді цист, одягнених щільними оболонками, або спор, а деякі мігрують на весь сухий період в сприятливіші за режимом вологості|вогкості| регіони. У|в,біля| мешканців|пожильців| вологих|вогких| місць – гігрофілів – пристосування до життя за цих умов не такі|настільки| різноманітні, як у|в,біля| мешканців|пожильців| сухих місць проживання — ксерофілів|. Це, напевно, пояснюється тим, що вологі|вогкі| місцепроживання значно менш екстремальні, ніж сухі.
У|в,біля| рослин, що мешкають за таких умов, як правило, відсутні пристосування проти|супроти| зав'ядання|в'янення| і швидкої втрати вологи, механічна тканина, як правило, розвинена слабо, сильно розвинені повітроносні| порожнини в корінні, стеблах|стеблинах| і листі. У|в,біля| тварин, пов'язаних з такими місцепроживаннями, спостерігається високий рівень водного обміну і відсутні пристосування до його регулювання.
Мезофільні рослини і тварини характеризуються рисами|межами|, проміжними між гігро-| і ксерофільними видами.
Правило Глогера, що полягає в тому, що підвиди того самого виду тварин, ссавців, що мешкають в більш аридних| умовах, забарвлені|пофарбовані| світліше за підвиди, властиві більш гумідним| районам, ґрунтується, напевно|певне|, на тому, що темніші речовини, які визначають забарвлення|фарбування| шерсті (меланіни), відкладаються інтенсивніше у вологіших|вогкіших| регіонах.
Опади, що випадають на суші, мають різну долю: вони можуть утворювати поверхневий|поверховий,зверхній| і внутрішньо ґрунтовий|внутрішньоґрунтовий| стік, випаровуватися, транспіруватися рослинами. На всі перераховані компоненти рослинний покрив чинить той чи інший вплив. Ліс самим своїм існуванням сприяє інтенсивнішому випаданню опадів, над відкритими|відчиненими| просторами їх випадає менше. Частка|доля| опадів, що затримується кронами дерев в лісі або травостаном| на луках, у преріях і саванах, різна і залежить від особливостей рослинного покриву. Так, в різних типах лісу частка|доля| опадів, що затримуються кронами, стікаючих по стовбурах|стволах| дерев і проникаючих|проникних| крізь крони безпосередньо до ґрунту, різна. Вплив на затримання опадів чагарникових і трав'янистих угруповань значно слабший у порівнянні з лісом, але|та| виражений|виказано,висловлено| досить чітко. У лісі зростає відносна вологість|вогкість| повітря в порівнянні з відкритими|відчиненими| ділянками, зменшується випаровування з поверхні ґрунту (чому сприяє лісова підстилка). Проте|однак| до вологи, що випаровується з поверхні ґрунту, в угрупованнях додається і волога, що випаровується з поверхні стовбурів|стволів|, стебел|стеблин| і листя. Ліс може повернути в атмосферу до 1/3 вологи, що випала над ним у вигляді опадів. У трав'янистих угрупованнях ця частка|доля| менша.
Транспірація, здійснювана лісовими рослинами, значно інтенсивніша транспірації лугових трав. Для заліснених територій характерне різке уповільнення поверхневого|поверхового,зверхнього| стоку, який в цих випадках не має катастрофічного характеру|вдачі|.
Вітер відіграє істотну|суттєву| роль в розповсюдженні|поширенні| насіння і плодів, спор рослин і пасивних зачатків тварин. Плоди і насіння рослин можуть мати спеціальні пристосування, що полегшують це перенесення та|перенос| що збільшують парусність| переносимих зачатків. Це або летючки|летучки| з|із| більш менш численних|багаточисельних| волосків, або крилатки, плівчасті або шкірясті. Інколи плодик вміщений|ув'язнений| в плівчастий мішечок, що також полегшує його перенесення|перенос|. Не менше значення має вітер і для запилення квіток рослин. Квітки вітрозапилюваних рослин зазвичай виробляють значно більшу кількість пилку в порівнянні з запилюваними комахами, оскільки|тому що| в першому випадку шансів потрапити|попасти| на рильце маточки у|в,біля| порошинок багато менше. Порошинки легкі, іноді|інколи| з|із| повітряними мішками, що полегшують їх політ (наприклад, у|в,біля| хвойних). Рильця маточок вітрозапилюваних квіток висовуються з|із| квіток і часто бувають перистими|пір'ястими|, тому вони мають велику «уловлюючу поверхню». Вітрозапилювані рослини, як правило, ростуть|зростають| великими групами, багато дерев і чагарники з|із| квітками, що запилюються вітром, квітнуть до розпускання листя.
Таблиця 3
Річна витрата води в лісі і на лугу (у мм) (по В. Н. Оболенському, 1935)
| Рослинність | Випаровування з поверхні ґрунту | Транспірація | Всього |
| Ліс | |||
| Луг |
Одночасно вітер здійснює формоутворювальний вплив на рослини. Він може ламати гілки дерев, а вітер ураганної сили ламає біля основи стовбури|стволи| дерев або вириває їх з коренем. Так, в лісах вологих|вогких| тропіків утворюються прогалини на місці вирваних ураганом дерев. Величезні вітровальні ділянки спостерігаються в лісах помірного поясу, переважно хвойних, особливо ялинових, оскільки коріння ялини не йде глибоко в землю|ґрунт|, а простягається близько|поблизу| до поверхні ґрунту. Вітер сприяє утворенню прапороподібних| форм дерев, особливо біля верхньої межі|кордону| їх зростання: у горах і біля межі лісу на півночі. Поза сумнівом|безсумнівно|, що вітер відіграє істотну|суттєву| роль в безліссі| тундри. Механізм цього впливу оцінюється різними авторами по різному. А. О. Чильман вважає|гадає|, що взимку, коли ґрунт промерзає, а гілки, не укутані снігом, продовжують транспірувати, всі частини|частки| дерева, сніги, що підіймаються над поверхнею, висихають. Б. Н. Городков вважає|лічить|, що найбільш критичним періодом в цьому відношенні виявляється|опиняється| весна, коли гілки вже сильно прогріваються сонцем, а ґрунт з|із| корінням ще не відтанув. Якої з|із|цих точок зору не дотримуватися — значення вітру для безліссяо тундри поза сумнівом|безсумнівно|.
Проходження північно-східних меж|кордонів| ареалів широколистяних порід, властивих для Східної Європи|спілки,сполучника| (липа дрібнолиста зустрічається на північний схід лише до Тобольська, а східні межі|кордони| ареалів дуба звичайного і клена платаноподібного не доходять до Уралу), пов'язане з тим, що гілки цих порід взимку транспірують значно більше|значно більший| вологи на одиницю поверхні в порівнянні з хвойними, а низькі зимові температури при великій швидкості вітру не перешкоджають зимовій транспірації. У дослідах відомого російського ботаніка А. Я. Гордягина транспірація з одиниці поверхні гілок дуба звичайного й інших широколистяних порід в 2 рази і більш перевищує транспірацію з гілок модрини. При цьому транспірація гілок дуба не припинялася навіть при значних морозах: середні добові втрати ваги у|в,біля| цього виду складали при температурах 22 -8 і -18,4° відповідно 0,8, 0,6 і 0,2% живої|жвавої| ваги. А. Я. Гордягин указує|вказує|, що в березні такі втрати вологи не компенсуються за рахунок притоку|припливу| води із старіших частин|часток| дерева. Отже, інтенсивне зимове і ранньовесняне випаровування однорічними гілками дуба та інших широколистяних порід є|з'являється,являється| однією з причин, що перешкоджають їх поширенню в області із суворішим кліматом (на схід і північний схід). Роль вітру при цьому істотна|суттєва|.
При заселенні островів вітер грає подвійну|двоїсту| роль. По-перше, він сприяє перенесенню|переносу| на острови діаспор (насіння, плодів і спор) рослин, а також різних крилатих безхребетних тварин. Одночасно вітер відносить з островів і з узбережжя материків різні не дуже добре літаючі, хоч і крилаті форми. В результаті|унаслідок,внаслідок| відбору в таких місцепроживаннях багато видів, що мають крилатих родичів, що літають, стають безкрилими або, якщо зберігають крила, то набувають|придбавають| звички затаюватися під корою, серед листя і т.п. Отже, вітер проводить|виробляє,справляє| відбір серед мешканців|пожильців| ділянок, що зазнають дії вітру, сприяючи розвитку безкрилих форм.
Вплив лісових угруповань на швидкість вітру значний. Швидкість вітру, що дме у бік лісу, зменшується вже на деякій відстані від узлісся|опушення|. У лісі ж вона падає швидко. Аналогічний вплив, хоч і не настільки сильний, чинять на швидкість вітру й інші рослинні угруповання|спілки|.
Газовий склад атмосфери в природі різко змінюється лише навколо|навкруг,довкола| вулканів, де зростає вміст газів, шкідливих для життя тварин і рослин. На високогір’ї| зменшується загальний|спільний| тиск|тиснення| атмосфери і парціальний тиск|тиснення| кисню. У|в,біля| тварин здатність|здібність| пристосовуватися до пониженого вмісту кисню пов'язана із|із| збільшенням вмісту гемоглобіну в крові. Ті види, які мають значну пластичність цієї ознаки, можуть мешкати в горах вище за тварин, у|в,біля| яких вміст гемоглобіну в крові змінюється у вузьких межах. У|в,біля| рослин можливість|спроможність| існувати на високогір’ї| пов'язана з підвищеним вмістом хлорофілу в тканинах листу|аркуша|, проте|однак| механізм цього впливу не цілком|сповна| зрозуілий. Для оцінки і наочного|наглядного| уявлення про вплив основних параметрів клімату на життя організмів застосовується метод клімограм| (кліматограм), при якому на осі координат наносяться|завдаються| основні кліматичні чинники|фактори| (температура і вологість|вогкість| повітря або температура і опади). Є|наявний| різні способи побудови|шикування| клімограм| (мал. 21). Особливо детально розроблені способи побудови|шикування| клімограм| німецьким ботаніком Г. Вальтером. На них показане не лише співвідношення температур і опадів, але й тривалість сухого і вологого|вогкого| періодів і деякі інші показники.
ЕДАФІЧНІ ЧИННИКИ|фактори|
Едафічними чинниками|факторами| є|з'являються,являються| особливості ґрунтів і снігового покриву|покривала|. При цьому важливі|поважні| хімічні властивості субстрату (концентрація водневих іонів, що визначає кислотність ґрунту, вміст легкорозчинних солей|соль| і поживних речовин тощо) і його фізичні особливості (механічний склад і рихлість, ступінь|міра| аерації тощо).
Кислотність ґрунту, що виражається|виказується,висловлюється| концентрацією водневих іонів рН| (при рН| 7 реакція нейтральна, більше 7 – лужна, менше 7 – кисла), суттєва для тварин і особливо для рослин. Кожен вид рослини існує при певній амплітуді рН| і має свій оптимум цієї величини. Так, сильнокислим| ґрунтам (рН| 3,5) відповідає оптимум сфагнових мохів, рН| 4 – оптимум щупака звивистого і хрестовника| лісового, рН| 6-7 – мати-й-мачухи і ячменю дворядного тощо. Різні види найпростіших мешкають при рН| 3,9-9,7; серед них є|наявний| види, що надають перевагу кислим, нейтральним і лужним ґрунтам. Дощові черв'яки не переносять рН| нижче 4,4. Наземні молюски, що мають мушлі, для побудови|шикування| яких потрібен кальцій, живуть при рН| 7-8.
З|із| хімічних елементів, що отримуються рослинами з|із| ґрунту, найбільше екологічне значення мають азот, фосфор і калій. Вони споживаються в значних кількостях і тому їх не вистачає частіше за інших.
За змістом основних поживних речовин ґрунти можуть бути бідними, середніми і багатими. Відповідно рослини, що живуть на цих ґрунтах, — оліготрофними, мезотрофними і евтрофними. Деякі рослини можуть бути названі|накликати| нітратними, вони зазвичай добре розвинені на ґрунтах, багатих на солі азоту. Такими є пирій повзучий, пшениця, кращі кормові злаки, хміль, кропива дводомна, іван-чай, вільха і деякі інші. Вміст в ґрунті солей|соль| кальцію сприяє створенню|створінню| лужної реакції ґрунтів, воно сприяє обміну речовин у|в,біля| вищих рослин. По відношенню до кальцію всі рослини поділяються на кальцієфоби| (що уникають кальцію) і кальцієфіли| (пов’язані з багатими на кальцій ґрунтами). Є|наявний| і види, байдужі до кальцію. У флорі центральної частини|частки| Східної Європи до кальцієфілів| належать багато орхідей, костер річковий, люцерна серповидна та інші, з|із| дерев — модрина сибірська, бук і ясен. Кальцієфобами є|з'являються,являються| сфагнові мохи і рослини, пов'язані зі сфагновими болотами, а також щавель кислий, види щупака, з|із| дерев — справжній|теперішній,даний| каштан, з|із| чагарників — чай. Не виключено, що пов’язаність рослин з ґрунтами з|із| різним вмістом кальцію пов'язана із ступенем|мірою| їх кислотності. Серед тварин є|наявний| також види, приурочені до ґрунтів з|із| високим вмістом кальцію: з|із| червів Allolobophora caliginosa, Eisenia rosea, E. terrestris, з|із| амеб, раковин Centropyxis plagistoma, Geopyxella sylvicola; черв Allolobophora icterica мешкає на ґрунтах, бідних на кальцій.
З ґрунтами, багатими на легкорозчинні солі|соль|, пов’язані рослини, що називаються галофітами|. Вони характеризуються дуже високим осмотичним тиском|тисненням|, що досягає 50-100 атм|, часто мають сукулентну| будову|споруду|, нерідко|незрідка| повністю або майже повністю позбавлені листя, накопичують|скупчують,нагромаджують| від 25 до 84% зольного залишку|остачі|. Ці особливості дають їм можливість|спроможність| існувати там, де вміст легкорозчинних солей|соль| настільки великий, що вони нерідко|незрідка| утворюють суцільний потужний шар на поверхні ґрунту.
Із|із| засоленими ґрунтами пов'язані й деякі тварини. Лише в Середній Європі до солонців і солончаків приурочено близько 20 специфічних родів жуків, що зустрічаються виключно|винятково| або майже виключно|винятково| на цих ґрунтах.
Фізична будова|споруда| ґрунтів також чинить дуже суттєвий вплив на рослин і тварин, що мешкають на них і в них. Піщані ґрунти різко відрізняються від ґрунтів суглинистих і глинистих. Пісок — рухливий субстрат, що має рихлу структуру. Мешканці|пожильці| пісків (псамофіли|) як рослини, так і тварини мають специфічні особливості, що дозволяють їм використовувати цей субстрат. Рослини пісків здатні|здібні| давати додаткове коріння при засипанні нижніх частин|часток| їхніх стебел|стеблин| і переносити оголення кореневих систем при перевіванні пісків на інше місце. У|в,біля| багатьох з|із| них є|наявний| довгі кореневища, що скріплюють субстрат. Коріння псамофілів| одягнене чохликами| з|із| часток піску, що надійно захищають їх від висихання. Такі чохлики| пов’язані з корінням тільки|лише| в місцях розгалуження останніх або там, де від них відгалужуються дрібні|мілкі| корінці. Зсередини чохлики| вистелені корою, що раніше вкривала корінці. Листя псамофілів| жорстке, вузьке, а у|в,біля| багатьох видів цілком відсутнє, що полегшує опір вітру, що нерідко|незрідка| несе хмари піску. Плоди псамофілів| одягнені плівчастою оболонкою, що утворює міхур|пузир,пухир|, або становлять кульки з|із| пружними волосками на поверхні. Вони добре пристосовані до перекочування по піску.
Тварини, що мешкають в пісках, мають здатність|здібністю| швидко в них закопуватись. Ящірки-круглоголовки закопуються з|із| такою швидкістю|прудкістю| і так добре і швидко зарівнюють те місце, де вони закопались, що, навіть спостерігаючи за цим процесом, важко|скрутно| виявити, де ж під рівною поверхнею піску затаїлась ця тварина. На лапках багатьох псамофільних тварин є|наявний| щіточки з|із| волосків або рогові луски, що збільшують їх поверхню. Це пристосування для успішнішого риття нір в рихлому ґрунті, а також, напевно, для швидкого пересування по піску. Але|та|, незважаючи на ці пристосування, нори в піску виходять дуже неміцні і недовговічні. Тому мешканці|пожильці| пісків не будують глибоких нір на тривалий час. Багато мешканців|пожильців| піску (змії, ящірки, ряд|лава,низка| комах) можуть пересуватися в товщі піску на значні відстані.
Значення снігового покриву|покривала| в житті рослин і тварин різноманітне. Він вкриває|укриває| поверхню ґрунту і тим самим сприяє збереженню|зберіганню| тепла в ґрунті. У багатосніжних районах можливий підсніжний розвиток рослин, які після|потім| цього зацвітають ранньою весною. Тварин по відношенню до сніжного|снігового| покриву|покривала| А. Н. Формозов запропонував поділити на дві групи: хіонофобів| — «снігоненавистників|», для яких сніговий покрив|покривало| несприятливий, оскільки|тому що| вони ведуть надсніжний| спосіб життя, і хіонофілів| — «сніголюбів|», які мешкають під снігом і для яких сніговий покрив|покривало| створює сприятливіші умови існування. Малосніжні зими виявляються|опиняються| несприятливими для «хіонофілів|» (кротів, полівок), які гинуть в ці зими масами, але зате виявляються|опиняються| сприятливими для «хіонофобів|» (копитних тварин), яким легше пересуватися по малопотужному сніговому покриву|покривалу|, а також для мишей тощо. Багатосніжні зими чинять протилежний вплив. Для тварин, що пересуваються по снігу, велику роль відіграє навантаження на підошви лап, пов’язане з вагою тварини і поверхнею лап. Вагове навантаження на 1 см2 поверхні лап складає у|в,біля| кабана 903 г, лося – близько 500, вовка – 190, дикого північного оленя – 140, зайців – 19-24, горностая – 8-10, хатньої миші – 5,5, самця глухаря – 59 і білої куріпки – 12 г. До зими опорна поверхня багатьох тварин збільшується за рахунок волосся, щетин, пір’я, рогових щитків. Для таких довгоногих звірів, як лось, сніг завглибшки 50-40 см не є|з'являється,являється| перешкодою до швидкого пересування. Вагове навантаження у 10-15 г на 1 см2 спостерігається у|в,біля| видів, що швидко пересуваються по снігу; при навантаженні 30-40 г на 1 см2 коротконогі звірі насилу пересуваються по рихлому снігу.
Багато тварин при випадінні снігу мігрують. Утруднюється не лише пересування по снігу, але й пошуки тваринами корму, укритого снігом. Там, де на снігу утворюється кірка|скоринка|, корм стає недоступним або малодоступним. У|в,біля| осілих звірів і птахів|птиць| взимку спостерігається перехід на інші корми. Так, зайці взимку харчуються значною мірою гілковим кормом, а влітку – травами.
Дрібні|мілкі| тварини, що ведуть підсніжний спосіб життя, залишаються взимку в умовах глибокого снігового покриву|покривала| активними. Деякі тварини, наприклад тетеревині птахи|птиці|, використовують сніговий покрив|покривало| як тимчасовий притулок, влаштовуючи|владнуючи,улаштовувавши| в ньому нори на ніч.
Вологість|вогкість| ґрунту справляє істотний|суттєвий| вплив на поширення організмів. Ґрунт може бути фізично сухим, вологим|вогким| тією чи іншою мірою, і фізіологічно сухим. Фізіологічно сухим ґрунт може бути під впливом різних причин. Цей термін застосовують до ґрунту, що містить|утримує| вологу удосталь, але|та| в такому стані, що вона не може бути використана рослинами і якоюсь мірою тваринами. Так, фізіологічно сухими є засолені ґрунти, в яких волога завдяки високому осмотичному тиску|тисненню| розчинів солей|соль| не може або майже не може бути добута рослинами. Фізіологічно сухими є|з'являються,являються| багаті на вологу, але такі що мають дуже низьку температуру, ґрунти тундри. Фізіологічно сухі місцепроживання екстремальні, і їхні мешканці|пожильці| повинні бути пристосовані або до різко підвищеного вмісту в ґрунтовому розчині легкорозчинних солей|соль|, або до незвично низьких температур ґрунтового середовища|середи|.
У верхових сфагнових болотах фізіологічна сухість при майже повному|цілковитому| насиченні мохового покриву|покривала| водою пов’язана з високим вмістом у воді розчинених високомолекулярних сполук, у меншій мірі — з|із| низькими температурами води.
Як вже було сказано, гігрофілами| називають види, що мешкають на вогких ґрунтах, гідрофітами| — мешканців|пожильців| водного середовища|середи|.
ГІДРОЛОГІЧНІ ЧИННИКИ|фактори|
Специфіка водного середовища|середи| полягає вїї значно більшої щільності в порівнянні з повітряною, в меншій прозорості, в наявності течій, в різноманітності солоності, в різкому підвищенні тиску|тиснення| і пониженні освітленості з|із| глибиною.
Вода має рівніший хід температур в порівнянні з сушею. По-перше, її температура не може падати скільки-небудь помітно нижче нуля без замерзання. По-друге, зміни температури від місця до місця відбуваються|походять| у водоймищах досить поступово; по-третє, завдяки значній теплоємності води, її нагрівання і охолоджування відбуваються|походять| при однаковій інтенсивності сонячних променів значно повільніше в порівнянні з сушею. У тропічному поясі температура води постійно висока, в Арктиці і Антарктиці — постійно низька, в помірних широтах коливання температури води найбільші, та все ж вони значно менші, ніж на суші. Водні організми, як правило, стенотермні|.
Зважаючи на|внаслідок,унаслідок| значні відмінності|відзнаки| умов водного середовища|середи| від умов, в яких протікає життя наземних організмів, водні організми будуть розглядаються окремо|розгледіти|, в розділі, присвяченому життю мешканців|пожильців| вод.
ОРОГРАФІЧНІ ЧИННИКИ|фактори|
Орографічні чинники|фактори|, на відміну від едафічних, з|із| якими їх раніше об'єднували в одну групу, чинять на організми не лише прямий, але і непрямий вплив. Розташування на тому чи іншому елементі макрорельєфу (схилі певної експозиції і крутизни|крутості|, певній висоті над рівнем моря) справляє істотний вплив на умови існування організму. Ця сторона впливу рельєфу буде детально розглянута у розділі|на чолі|, присвяченому значенню висотної поясної в поширенні організмів. Але|та|, окрім|крім| цієї прямої дії, макрорельєф чинить вплив і на характер|вдачу| органічного світу, перегороджує шлях|колію,дорогу| вітрам, що мають певні властивості (наприклад, що несуть атмосферні маси, багаті на вологу або, навпаки, сухі). Такий вплив може поширюватися дуже далеко від місця розташування гірської|гірничої| країни. Так, північноамериканські Кордільєри, що знаходяться|перебувають| на шляху|колії,дорозі| вологої|вогкої| західної вітрової циркуляції, є|з'являються,являються| основною причиною того, що зони рослинності в Північній Америці, починаючи|розпочинаючи,зачинаючи| з|із| широколистяних лісів, розташовуються не з півночі на південь, а зі сходу на захід. У напрямі|направленню| зі сходу на захід змінюють один одного в цій країні широколистяні ліси, високотравні і низькотравні| прерії, напівпустелі і пустелі. Кавказький хребет, що перегороджує шлях|колію,дорогу| на південь холодним вітрам, робить|чинить| можливим існування в Закавказзі субтропіків тощо.
Окрім|крім| макрорельєфу, в житті організмів істотну|суттєву| роль відіграє мезо-| і мікрорельєф. Мезорельєф позначається на розподілі зональних, інтра-| і екстразональних угруповань в межах географічних зон. Мікрорельєф впливає на перерозподіл солей|соль| і вологи, сприяючи створенню|створінню| комплексів угруповань |спілок|, тобто чергуванню невеликих за розмірами ділянок угруповань |спілок|, пов’язаних з підвищеними і пониженими ділянками мікрорельєфу.
Враховуючи різноманітний вплив рельєфу на життя тварин і рослин, орографічний чинник|фактор| винесений в особливу групу екологічних чинників|факторів|.
БІОТИЧНІ| ЧИННИКИ|фактори|
Організми, мешкаючи спільно, вступають один з|із| одним в різні відносини. Ці відносини отримали назву біотичних|. Ми можемо класифікувати біотичні| відносини за тим, як вони позначаються на компонентах, що вступають в ці відносини. Якщо позначити позитивні результати відносин для одного з пари організмів, що знаходяться|перебувають| в цих відносинах, знаком « + », негативні|заперечні| результати знаком «—», а відсутність як позитивних, так і негативних|заперечних| результатів знаком «0», то ми отримаємо таку таблицю типів відносин між організмами.
Типи біотичних відносин
| Результат для організму А | Результат для організму Б | Найменування типу відносин | |
| _ | |||
| + | + | мутуалізм коменсалізм | симбіоз |
| + | — | Хижацтво, паразитизм | |
| + | нейтралізм | ||
| — | антибіоз | ||
| — | — | конкуренція |
Ця дещо схематична класифікація дає загальне|спільне| уявлення про різноманітність біотичних| відносин. Наведемо приклади|зразки| відносин різних типів.
Прикладом|зразком| мутуалістичних| відносин є|з'являються,являються| відносини між раком-самітником, що ховає м'яке черевце в порожню|пусту| мушлю молюска, і актинією, що живе на цій мушлі. Виростаючи, рак-самітник замінює раніше зайняту|позичену,посісти| ним мушлю на більшу. При цьому він обережно знімає актинію із старої мушлі і пересаджує її на нову. Актинія, що має жалючі клітини, дає раку-самітнику додатковий захист від ворогів, сама ж, пересуваючись на мушлі, зайнятій|позиченій,посісти| раком-самітником, опиняється в кращих умовах харчування, поступово змінюючи місцеположення разом з твариною, що повільно переповзає по дну. У мурашниках нерідко|незрідка| поселяються деякі жуки, яких мурашки охороняють і годують, в свою чергу злизуючи виділення із|із| залозок жуків. Так само мурашки захищають, тлю|попелюхою|, що смокче|ссуть| рослинні соки, іноді|інколи| навіть зводять|підносять| над ними подобу галерей, а самі харчуються солодкими виділеннями тлі|попелюхою|.
Коли нільський крокодил лежить на березі, невелика пташка трохілус| може забиратися до нього в пащу і вибирати там п'явок що присмоктались і застряглі між зубів залишки їжі. Взаємна вигода таких відносин ясна.
Зелені водорості мешкають в колоніях коралових поліпів — будівників рифів. Вони асимілюють, доставляючи коралам органічну речовину і в свою чергу отримують від поліпів захист від ворогів. Різні зелені водорості поселяються в клітинах одноклітинних найпростіших. Водорості захищені від несприятливих впливів тілами їх господарів|хазяїв|, а прозорість цих тіл дозволяє водоростям асимілювати, забезпечуючи господарів|хазяїв| вуглеводами і одержуючи|отримуючи| від них азотисті сполуки і вуглекислоту. У такі самі відносини із|із| зеленими водоростями вступають багато губок, деякі черви, голкошкірі й молюски. Іноді|інколи|, як у|в,біля| черва Convoluta, ці відносини настільки тісні, що ні черв ні водорості, що мешкають в ньому, не можуть існувати самостійно.
Бактерії й напростіші, розкладаючі клітковину, поселяються в кишечнику як безхребетних (терміти), так і деяких хребетних|хребцевих| (жуйні ссавці та ін.) тварин, переводячи|перекладаючи,переказуючи| клітковину в доступні для засвоєння господарями|хазяями| форми і одержуючи|отримуючи| в свою чергу від господарів|хазяїв| захист і харчування.
Складні відносини між хижими мурахами і так званими мірмекофільними (мурахолюбними|) рослинами. Мурахи поселяються в особливих порожнинах, наявних в стовбурах|стволах| мірмекофільних дерев. Нерідко|незрідка| рослини-господарі|хазяї| виробляють спеціальні високоживильні|живлячі,поживні| тільця, багаті на білки, протеїди й інші харчові речовини, названі тільцями Белта і Мюллера, якими харчуються мурахи-квартиранти. Ці хижі мурахи захищають рослин-господарів|хазяї| від мурахів-листогризів| й інших рослиноїдних комах, які, якщо рослина не має захисників, нерідко|незрідка| за короткий час повністю об'їдають листя і роблять|чинять| його існування неможливим.
До мутуалістичних| відносин належить також участь тварин в запиленні рослин і перенесенні|переносі| плодів і насіння. До цього ж типу відносин слід віднести і взаємостосунки клубенькових| бактерій з|із| рослинами-господарями|хазяями|, взаємостосунки грибів, що створюють мікоризу на корінні багатьох рослин, з|із| рослинами-господарями|хазяями|.
Перехресне запилення рослин призводить|призводить,наводить| до створення|створіння| потомства з|із| кращими спадковими|спадкоємними| властивостями, ніж ті, які виникають в результаті|унаслідок,внаслідок| самозапилення. Тому взаємні пристосування рослин і обпилювачів|обпильників|, якими є|з'являються,являються| переважно комахи, рідше деякі птахи|птиці| і кажани, дуже складні і різноманітні. При перенесенні|переносі| діаспор (плодів і насіння) тварини одержують|отримують| додаткове харчування.
Взаємовідношення клубенькових| бактерій, а також грибів що створюють мікоризу з|із| господарями|хазяями| нерідко|незрідка| такі|настільки| складні, що їх іноді|інколи| відносять до взаємного паразитизму, при якому спочатку бактерія або гриб паразитує на господарі|хазяїні|, а потім господар|хазяїн| паразитує на бактерії або грибі. Такі|настільки| ж складні, хоча в ще більшій мірі наближаються до мутуалістичних|, відносини між грибом і водорістю в лишайнику, в результаті|унаслідок,внаслідок| яких виникли лишайники, що можуть існувати в таких екстремальних умовах, в яких ні гриб, ні водорість жити не можуть.
До відносин коменсалізму відносяться взаємостосунки між рибами-прилипалами та їх господарями|хазяями|. Прилипало спинним плавником, перетвореним на присосок, прикріпляється до шкіри акули та інших великих риб, переноситься ними на значні відстані й харчується залишками їжі цих риб. Приблизно такі самі відносини існують між рибами і що прикріпляються до них гідроїдними| поліпами, між китоподібними і сидячими на них вусоногими| раками, так званими «морськими жолудями».
Черв'як з|із| роду нереїс| Nereis, що має невеликі розміри, поселяється усередині мушлі, зайнятої|позиченої,посісти| раком-самітником, і поїдає залишки його їжі.
Інфузорії-опалини поселяються в задній кишці жаби, де харчуються напівперевареною їжею господарів|хазяїв|.
Нерідко|незрідка| протекційне забарвлення|фарбування| і форма, властиві тваринам, роблять|чинять| їх малопомітними саме на тлі|на фоні| рослин, з|із| якими вони схожі, іноді|інколи| ж більш-менш рідкісна|рідка| нешкідлива комаха наслідує рясній|багатій| отруйній формі (псевдоапосематична| форма або забарвлення|фарбування|), що запобігає нападу на нього ворогів. Це також приклад|зразок| прояву|вияву| коменсалізму.
Водорості, що поселяються| на поверхні тіла, в шерсті тварин, роблять|чинять| цих тварин менш помітними на зеленому фоні листя. Особливо незвичайна|незвична| в цьому відношенні зелена водорость, що мешкає в довгих жолобчастих волосках південноамериканського ссавця — лінивця. Іноді|інколи| вона розвивається в таких масах, що надає тварині зелене забарвлення|фарбування|, яке служить додатковим захистом від ворогів.
Тварини дуже часто одержують|отримують| укриття, поселяючись| в дуплах рослин, під їх корою, що теж|також| є формою коменсалізму.
До коменсалів| належать епіфіти|, що представляють|уявляють| групу рослин, особливо широко розвинену у вологих|вогких| тропічних лісах. Вони можуть бути поділені на чотири групи. Епіфіти| з|із| цистернами, поширені тільки|лише| в тропічній Америці, що належать до родини бромелієвих|, утворюють розетки листя, в яких збирається вода. У таких мініатюрних, піднятих на гілки дерев, ставках мешкають водні рачки, личинки комарів, пуголовки жаб й іноді|інколи| навіть комахоїдні рослини — пухирчатки. Для епіфіти|-бра і гніздових епіфітів| характерні|вдача| сплетення повітряного коріння; ґрунт, який утримується|стримується| серед них, дуже багатий на гумус (нерідко|незрідка| його більше 40%). Напівепіфіти|, на відміну від епіфітів| інших типів, не втратили повністю зв'язків із ґрунтом, в який опускають з гілок дерев при|даткові| коріння. Проте|однак|, якщо це коріння перерізати, це не перешкодить нормальному розвитку напівепіфітів|. Це ніби епіфіти| у момент становлення. До них належать багато видів родини ароїдних|. Нарешті|урешті|, протоепіфіти|, до яких належать усі решта епіфіти|, що поселяються| на стовбурах|стволах| і гілках дерев, не створюють ні запасів води, ні високогумусованого| ґрунту і живуть за рахунок тих органічних залишків, які є|наявний| на корі дерев.
Епіфіти|, які поселяються на листі (лише в тропічних і екваторіальних лісах, де тривалість життя листа|аркуша| вимірюється роком і більше), називаються епіфілами. Одні з епіфітів| світлолюбні і посухостійкі і мешкають на освітлених ділянках стовбурів|стволів|, інші — тінелюбні| й вологолюбні — на затінених ділянках стовбурів|стволів|.
Окрім|крім| епіфітів|, до коменсалім| належать ліани, які потребують опори, щоб піднятися|підійнятися| до світла. Ліан багато і в помірному поясі, але|та| вони переважно мають трав'янисті, нерідко|незрідка| однорічні стебла|стеблини|. У тропічних країнах багато ліан з|із| дерев'янистими стовбурами|стволами|. За способом підйому до світла ліани можуть бути поділені на декілька груп, одна з яких — ліани, що спинаються|видираються|, не мають жодних спеціальних пристосувань для лазіння на рослини, використовувані як опора. Іноді|інколи| у|в,біля| них розвиваються гілки, перпендикулярні до стовбура|стволу|, іноді|інколи| шпильки або колючки, що трохи полегшує опору. У|в,біля| в'юнких ліан стебло|стеблина| обвивається навколо|навкруг,довкола| опори. Ліани з|із| вусиками становлять найбагатшу на види групу ліан. Врешті, ліани, що лазять за допомогою коріння, розвивають повітряне коріння, яким чіпляються за опору.
Хижацтво в широкому сенсі|змісті,рації| слова — один з найбільш широко поширених типів біотичних| відносин. Консументи всіх порядків|ладів|, по суті, хижачать|хижакують|, споживаючи рослинну (консументи| першого порядку|ладу|) або тваринну (консументи| другого і третього порядків|ладів|) їжу. З погляду біотичних| відносин споживання|вжиток| живої|жвавої| рослинної маси не має принципових відмінностей|відзнак| від споживання|вжитку| тваринних кормів.
До відносин хижацтва примикає паразитизм. Паразитизмом називаються такі відносини, при яких один вид (паразит|шкідник|) харчується тканинами іншого виду (господаря|хазяїна|) і при цьому взаємостосунки паразита|шкідника| з|із| середовищем|середою| здійснюються в основному через організм господаря|хазяїна|. Грань між хижаком і паразитом|шкідником| провести нелегко.
Паразити можуть бути тимчасовими і постійними. Тимчасові паразити лише періодично нападають на господаря|хазяїна|, більш або менш т ривалий час харчуючись на ньому, постійні проводять на господарі|хазяїні| або в його тілі всю своє індивідуальне життя.
Переходи між постійними і тимчасовими паразитами поступові. Так, у|в,біля| овода паразитує лише личинка, доросла комаха веде вільний спосіб життя. Кліщі-аргазиди, що| мешкають в пустелях і напівпустелях, смокчуть|ссуть| кров по 4 хв. підряд, а іксодові| кліщі — до 8 діб|. Паразитичні черви (аскарида, волосоглав| і багато інших) все життя|усе життя|, окрім|крім| стадії яйця, проводять в тілі господаря|хазяїна|.
Паразитичні рослини — переважно постійні паразити, хоча у|в,біля| деяких з них, наприклад у|в,біля| спориньї|, є|наявний| фази розвитку, в даному випадку фаза склероція, який зимує поза|зовні| організмом господаря|хазяїна|, на поверхні ґрунту.
Є|наявний| паразити облігатні (обов’язкові) і факультативні (необов’язкові). Перші можуть вести тільки|лише| паразитичний спосіб життя, інакше гинуть, другі можуть бути паразитами, а можуть харчуватися сапрофітно, тобто мертвими рештками організмів.
За розташуванням паразита|шкідника| на або в тілі господаря|хазяїна| розрізняють паразитів зовнішніх і внутрішніх. Серед внутрішніх паразитів виділяють паразитів порожнинних, тканинних і внутрішньошкірних.
Серед рослин розрізняють повних|цілковитих| паразитів і напівпаразитів. Перші (наприклад, вовчок, повилика|) все харчування отримують від господарів|хазяїв|. Інші (омела, погремки|, очанки| та багатоінших|) фотосинтезують| самостійно, а від господарів|хазяїв| беруть тільки|лише| розчини мінеральних речовин.
Паразити можуть бути специфічними або неспецифічними. Перші мають одного господаря|хазяїна| або пов'язані з певною серією з|із| двох—трьох| послідовно змінюючих один одного господарів|хазяїв|. Інші можуть вражати|приголомшувати| широке коло рослин або тварин. Так, аскарида людини або споринья| іржі – приклади|зразки| специфічних паразитів, а іксодовий| кліщ або борошниста роса — неспецифічні паразити. Врешті, бувають паразити одногосподарні|, двогосподарні| й трьохгосподарні|. Перші в циклі свого розвитку вражають|приголомшують| лише один організм, в якому проходять|минають,спливають| всі стадії розвитку. Такою є, наприклад, аскарида. Двогосподарні паразити змінюють в циклі розвитку двох господарів|хазяїв|, трьогосподарні| — трьох. Приклади|зразки| двохгосподарних| паразитів — іржавинні гриби, першим з|із| господарів|хазяїв| яких є|з'являється,являється| барбарис або (у|в,біля| іншого виду гриба) — жостір|крушина|, а другим — злаки (пшениця, жито або овес|вівсюг|). Двогосподарним є|з'являється,являється| збудник малярії — плазмодій. Трьохгосподарний паразит|шкідник| — котяча двоус
Читайте також:
- IV. Закріплення й узагальнення знань
- IV. УЗАГАЛЬНЕННЯ І СИСТЕМАТИЗАЦІЯ ВИВЧЕНОГО
- Tип особистості й уявлення про самого себе.
- V. Систематизація і узагальнення нових знань, умінь і навичок
- V. Узагальнення та систематизація знань учнів.
- V. Узагальнення та систематизація знань учнів.
- V.УЗАГАЛЬНЕНН І СИСТЕМАТИЗАЦІЯ НОВИХ ЗНАНЬ І УМІНЬ УЧНІВ.
- V.УЗАГАЛЬНЕНН І СИСТЕМАТИЗАЦІЯ НОВИХ ЗНАНЬ І УМІНЬ УЧНІВ.
- VI. Узагальнення та систематизація знань
- VI. Узагальнення та систематизація знань учнів.
- VI. Узагальнення та систематизація знань.
- VII. Узагальнення та систематизація знань учнів.
| <== попередня сторінка | | | наступна сторінка ==> |
| | | ІНДИКАЦІЯ |
|
Не знайшли потрібну інформацію? Скористайтесь пошуком google: |
© studopedia.com.ua При використанні або копіюванні матеріалів пряме посилання на сайт обов'язкове. |